Abstract
Research efforts to account for elevated risk behavior among adolescents have arrived at an exciting new stage. Moving beyond laboratory studies of age differences in risk perception and reasoning, new approaches have shifted their focus to the influence of social and emotional factors on adolescent decision making. We review recent research suggesting that adolescent risk-taking propensity derives in part from a maturational gap between early adolescent remodeling of the brain’s socioemotional reward system and a gradual, prolonged strengthening of the cognitive-control system. Research has suggested that in adolescence, a time when individuals spend an increasing amount of time with their peers, peer-related stimuli may sensitize the reward system to respond to the reward value of risky behavior. As the cognitive-control system gradually matures over the course of the teenage years, adolescents grow in their capacity to coordinate affect and cognition and to exercise self-regulation, even in emotionally arousing situations. These capacities are reflected in gradual growth in the capacity to resist peer influence.
Introduction
"It seems like people accept you more if you’re, like, a dangerous driver or something. If there is a line of cars going down the road and the other lane is clear and you pass eight cars at once, everybody likes that. . . . If my friends are with me in the car, or if there are a lot of people in the line, I would do it, but if I’m by myself and I didn’t know anybody, then I wouldn’t do it. That’s no fun." — Anonymous teenager, as quoted in The Culture of Adolescent Risk-Taking (Lightfoot, 1997)
It is well established that adolescents are more likely than children or adults to take risks, as evinced by elevated rates of experimentation with alcohol, tobacco, and drugs, unprotected sexual activity, violent and nonviolent crime, and reckless driving (Steinberg, 2008). Early research efforts to identify the distinguishing cognitive immaturity underlying adolescents’ heightened risk-taking propensity bore little fruit. A litany of carefully controlled laboratory experiments contrasted adolescent and adult capacities to perceive and process fundamental components of risk information, but found that adolescents possess the knowledge, values, and processing efficiency to evaluate risky decisions as competently as adults do (Reyna & Farley, 2006).
If adolescents are so risk prone in the real world, why do they appear so risk averse in the lab? We propose that the answer to this question is nicely illustrated by the American teenager quoted above: “If I’m by myself and I didn’t know anybody, then I wouldn’t do it. That’s no fun.” If adolescents made all of their decisions involving drinking, driving, dalliances, and delinquency in the cool isolation of an experimenter’s testing room, those decisions would likely be as risk averse as those of adults. But therein lies the rub: Teenagers spend a remarkable amount of time in the company of other teenagers. In this article, we describe a new wave of research on the neurobehavioral substrates of adolescent decision making in peer contexts suggesting that the company of other teenagers fundamentally alters the calculus of adolescent risk taking.
Peer Influences on Adolescent Risk Behavior
Consistent with self-reports of lower resistance to peer influence among adolescents than among adults (Steinberg & Monahan, 2007), observational data point to peer influence as a primary contextual factor contributing to adolescents’ heightened tendency to make risky decisions. For instance, crime statistics indicate that adolescents typically commit delinquent acts in peer groups, whereas adults more frequently offend alone (Zimring, 1998). Furthermore, one of the strongest predictors of delinquent behavior in adolescence is affiliation with delinquent peers, an association that has been attributed in varying degrees to peer socialization (e.g., deviancy training; Dishion, Bullock, & Granic, 2002) and friendship choices, in which risk-taking adolescents naturally gravitate toward one another (e.g., Bauman & Ennett, 1996). Given the difficulty of distinguishing between these causal alternatives using correlational data, our lab has pursued a program of experimental research directly comparing the behavior of adolescents and adults when making decisions either alone or in the presence of peers.
In the first experimental study to examine age differences in the effect of peer context on risky decision making (Gardner & Steinberg, 2005), early adolescents (mean age = 14), late adolescents (mean age = 19), and adults (mean age = 37) were tested on a computerized driving task called the Chicken Game, which challenges the driver to advance a vehicle as far as possible on a driving course while avoiding crashing into a wall that could appear, without warning, on the course at any point. Peer context was manipulated by randomly assigning participants to play the game either alone or with two sameaged peers in the room. When tested alone, participants in each of the three age groups engaged in a comparable amount of risk taking. In contrast, early adolescents scored twice as high on an index of risky driving when tested with their peers in the room than when tested alone, whereas late adolescents’ driving was approximately 50% riskier in the presence of peers, and adults showed no difference in risky driving related to social context. The ongoing goal of our research program is to further specify the behavioral and neural mechanisms of this peer effect on adolescent risk taking.
A Neurodevelopmental Model of Peer Influences on Adolescent Decision Making
Building on extensive evidence demonstrating maturational changes in brain structure and function occurring across the second decade of life (and frequently beyond), we have advanced a neurodevelopmental account of heightened susceptibility to peer influence among adolescents (Albert & Steinberg, 2011; Steinberg, 2008). In brief, we propose that, among adolescents more than adults, the presence of peers “primes” a reward-sensitive motivational state that increases the subjective value of immediately available rewards and thereby increases preferences for the short-term benefits of risky choices over the long-term value of safe alternatives. Although a comprehensive presentation of the behavioral and neuroscientific evidence underlying this hypothesis is beyond our current scope (but see Albert & Steinberg, 2011), a briefreview of three fundamental assumptions of this model will set the stage for a description of our peer-influence studies.
First, decisions are a product of both cognitive and affective input, even when affect is unrelated to the choices under evaluation. Research with adult populations has identified several pathways by which affect influences decision making (Loewenstein, Weber, Hsee, & Welch, 2001). For instance, the anticipated emotional outcome of a behavioral option—how one expects to feel after making a given choice—contributes to one’s cognitive assessment of its expected value. Indeed, affective states may influence decision processing even when the source of the affect is not directly related to the choices under evaluation. Such incidental affective influences are apparent in experiments demonstrating the effect of preexisting or experimentally elicited affective states on adult perception, memory, judgment, and behavior (Winkielman, Knutson, Paulus, & Trujillo, 2007).
One experiment illustrating this effect found that incidental positive emotion elicited via the presentation of masked happy faces caused participants to pour and drink more of an unfamiliar beverage than participants who had viewed angry faces, despite no differences in self-reported emotion between the two groups (Winkielman, Berridge, & Wilbarger, 2005). Consistent with evidence for extensive overlap in the neural circuitries implicated in the evaluation of socioemotional and choice-related incentive cues (e.g., frontostriatal circuitry, including ventral striatum and ventromedial prefrontal cortex; for a recent review, see Falk, Way, & Jasinska, 2012), Winkielman and his colleagues describe this priming effect as an instance of approach sensitization. That is, neural responses to positively valenced socioemotional stimuli—in this case, responses that do not even reach the level of conscious awareness—may sensitize approach responding to unrelated incentive cues. As we describe below, several characteristics of adolescent neurobehavioral functioning suggest that this approach-sensitization effect could be a particularly powerful influence on adolescent decision making in peer contexts.
Second, relative to adults, adolescents exhibit stronger “bottom-up” affective reactivity in response to socially relevant stimuli. Whereas some controversy remains regarding the degree to which adolescents are more or less sensitive than children and adults to nonsocial reward cues (Galvan, 2010; Spear, 2009), few scholars now dispute that adolescence is a period of peak neurobehavioral sensitivity to social stimuli (Burnett, Sebastian, Kadosh, & Blakemore, 2011; Somerville, 2013). Puberty-related increases in gonadal hormones have been linked to a proliferation of receptors for oxytocin within subcortical and limbic circuits, including the amygdala and striatum (Spear, 2009). Oxytocin neurotransmission has been implicated in a variety of social behaviors, including social bonding and heightened attention to positive social stimuli (Insel & Fernald, 2004). Along with concurrent changes in dopaminergic function within neural circuits broadly implicated in incentive processing (Spear, 2009), these pubertyrelated increases in gonadal hormones and oxytocin-receptor density contribute to changes in a constellation of social behaviors observed in adolescence.
Peer relations are never more salient than in adolescence. In addition to a puberty-related spike in interest in opposite-sex relationships, adolescents spend more time than children or adults interacting with peers, report the highest degree of happiness in peer contexts, and assign the greatest priority to peer norms for behavior (Brown & Larson, 2009). This developmental peak in affiliation motivation appears to be highly conserved across species: Adolescent rats also spend more time interacting with peers than do younger or older rats, while showing evidence that such interactions are highly rewarding (Doremus-Fitzwater, Varlinskaya, & Spear, 2010). Moreover, several developmental neuroimaging studies have indicated that, relative to children and adults, adolescents show heightened neural activation in response to a variety of social stimuli, such as facial expressions and social feedback (Burnett et al., 2011). For instance, one of the first longitudinal neuroimaging studies of early adolescence demonstrated a significant increase from ages 10 to 13 in ventral striatal and ventral prefrontal reactivity to facial stimuli (Pfeifer et al., 2011). Together, this evidence for hypersensitivity to social stimuli suggests that adolescents may be more likely than adults to experience heightened approach motivation when exposed to positively valenced peer stimuli in decision-making scenarios, thus setting the stage for an exaggerated approachsensitization effect of peer context on decision making.
Third, adolescents are less capable than adults of “topdown” cognitive control of impulsive behavior. In contrast to social processing, which undergoes relatively sudden changes around the time of puberty, cognitive capacities supporting efficient self-regulation mature in a gradual, linear pattern over the course of adolescence. In parallel with structural brain changes thought to support neural-processing efficiency (e.g., increased axonal myelination), continued gains in response inhibition, planned problem solving, flexible rule use, impulse control, and future orientation occur during adolescence (Steinberg, 2008).
Indeed, evidence is growing for a direct link between structural and functional brain maturation during adolescence and concurrent improvements in cognitive control. In addition to studies correlating white-matter maturation with agerelated cognitive improvements (Schmithorst & Yuan, 2010), developmental neuroimaging studies using tasks requiring response inhibition (e.g., go-no/go, Stroop, flanker, and ocular-antisaccade tasks) have demonstrated relatively inefficient recruitment by adolescents of the core neural circuitry supporting cognitive control (e.g., lateral prefrontal and anterior cingulate cortex; Luna, Padmanabhan, & O’Hearn, 2010). Moreover, research on age differences in control-related network dynamics has demonstrated adolescent immaturity in the functional integration of neural signals deriving from specialized cortical and subcortical “hub” regions (Stevens, 2009). This immature capacity for functional integration may contribute to adolescents’ difficulties with simultaneously evaluating social, affective, and cognitive factors relevant to a given decision, particularly when social and emotional considerations are disproportionately salient.
Identification of Mechanisms Underlying Peer Influences on Adolescent Decision Making
In an effort to further delineate the neurodevelopmental vulnerability underlying adolescents’ susceptibility to peer influence, we have conducted a series of behavioral and neuroimaging experiments comparing adolescent and adult decision making in variable social contexts. Specifically, we have sought to determine whether the presence of peers biases adolescents’ decision making by (a) modulating responses to incentive cues, as predicted by the approach-sensitization hypothesis, (b) disrupting inhibitory control, or (c) altering both of these processes.
As a first step in addressing this question, we conducted an experiment in which we randomly assigned late adolescents (ages 18 and 19) to complete a series of tasks either alone or in the presence of two same-age, same-sex peers. Risk-taking propensity was assessed using the Stoplight game, a firstperson driving game in which participants must advance through a series of intersections to reach a finish line as quickly as possible to receive a monetary reward (Fig. 1). Each intersection is marked by a stoplight that turns yellow and, sometimes, red as the car approaches, and participants must decide to either hit the brakes (and lose time while waiting for the light to turn green) or run the light (and risk crashing while crossing an intersection). We also administered a go/no-go task as a measure of cognitive control and a delay-discounting task as a measure of preference for immediate over delayed rewards. Whereas no group differences were evident on the go/no-go index of inhibitory control, adolescents in the peerpresence condition took more risks in the Stoplight game (Albert et al., 2009) and indicated stronger preferences for immediate over delayed rewards (O’Brien, Albert, Chein, & Steinberg, 2011) than did adolescents who completed the tasks alone.
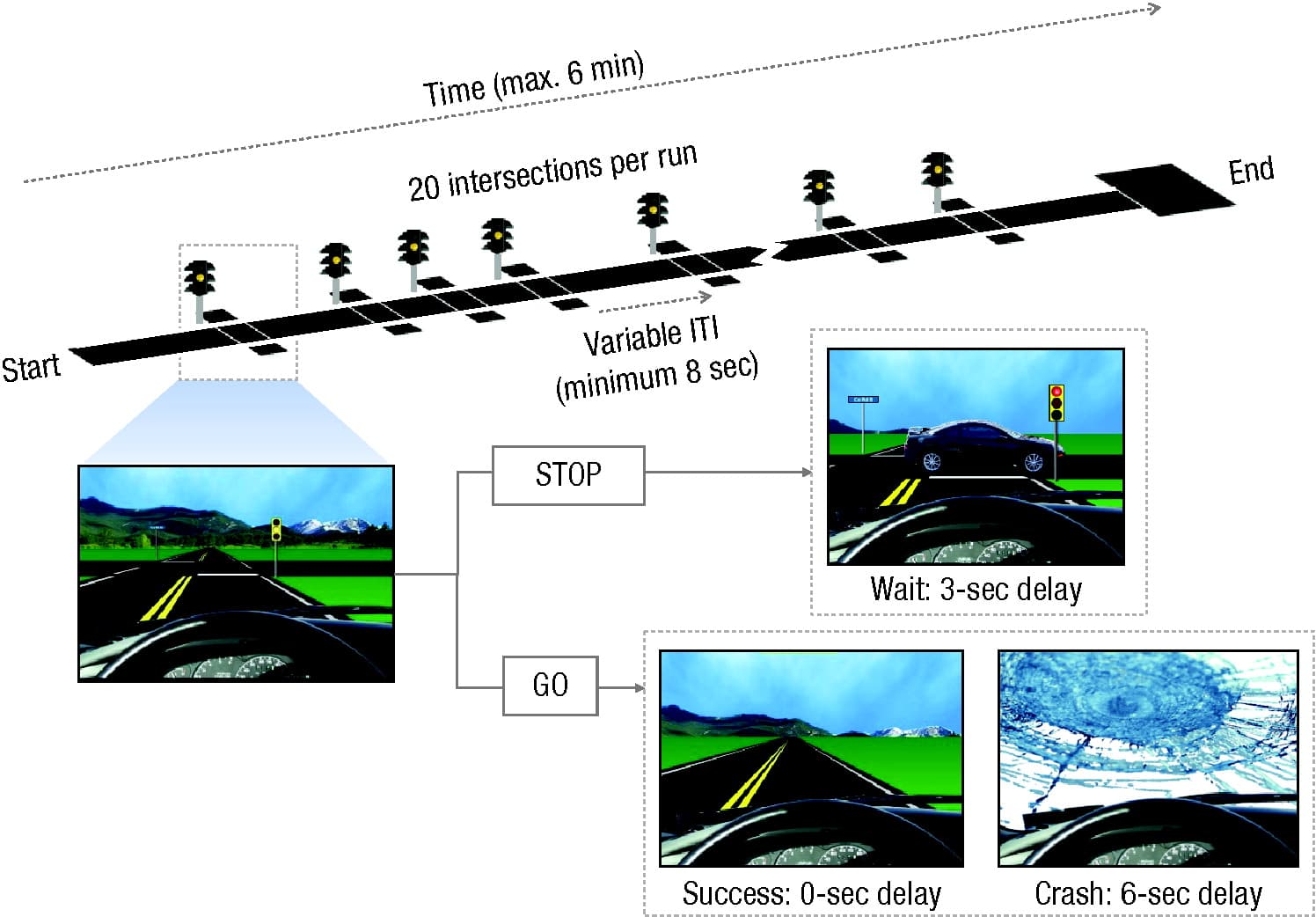
Fig. 1. Schematic of the Stoplight game. In this first-person driving game, participants are instructed to attempt to reach the end of a straight track as quickly as possible. At each of 20 intersections, participants must decide to either stop the vehicle (STOP) or take a risk and run the yellow or red light (GO). Stopping results in a short delay. Successful risk taking results in no delay, but unsuccessful risk taking results in a crash and a relatively long delay. Summary indices of risk taking include (a) the proportion of intersections at which the participant decides to run the light and (b) the total number of crashes. ITI = intertrial interval.
Findings from a recent follow-up experiment suggested that a peer’s observation influences adolescents’ decision making even when the peer is anonymous and not physically present. Using a counterbalanced repeated-measures design, we assessed late adolescents’ performance on a delaydiscounting task once in an alone condition and once in a deception condition that elicited the impression that the adolescents’ task performance was being observed by a same-age peer in an adjoining room. As predicted, participants exhibited a stronger preference for immediate rewards in the task when they believed they were being observed than when they were alone (Weigard, Chein, & Steinberg, 2011). Peer observation also increased rates of monetary gambles on a probabilistic gambling task, but only for participants with relatively low self-reported resistance to peer influence (Smith, Chein, & Steinberg, 2011). Along similar lines, Segalowitz et al. (2012) reported that individuals high in self-reported sensation seeking are particularly susceptible to the peer effect on risk tak-ing. Considered together, these behavioral results suggest that the presence of peers increases adolescents' risk taking by increasing the salience (or subjective value) of immediately available rewards, and that some adolescents are more susceptible to this effect than others.
Our recent work has used brain imaging to more directly examine the neural dynamics underlying adolescent susceptibility to peer influences. In the first of these studies, we scanned adolescents and adults while they played the Stoplight game, again using a counterbalanced within-subjects design (Chein, Albert, O'Brien, Uckert, & Steinberg, 2011).
All subjects played the game in the scanner twice once in an alone condition and once in a peer condition, in which participants were made aware that their performance was being observed on a monitor in a nearby room by two same-age, same-sex peers who had accompanied them to the experi-ment. As predicted, adolescents, but not adults, took significantly more risks when they were being observed by peers than when they were alone (Fig. 2). Furthermore, analysis of adolescents' neural activity during the decision-making epoch showed greater activation of brain structures implicated in reward valuation (ventral striatum and orbitofrontal cortex) in the peer-condition scans relative to the alone-condition scans, an effect that was not observed for adults (Fig. 3).
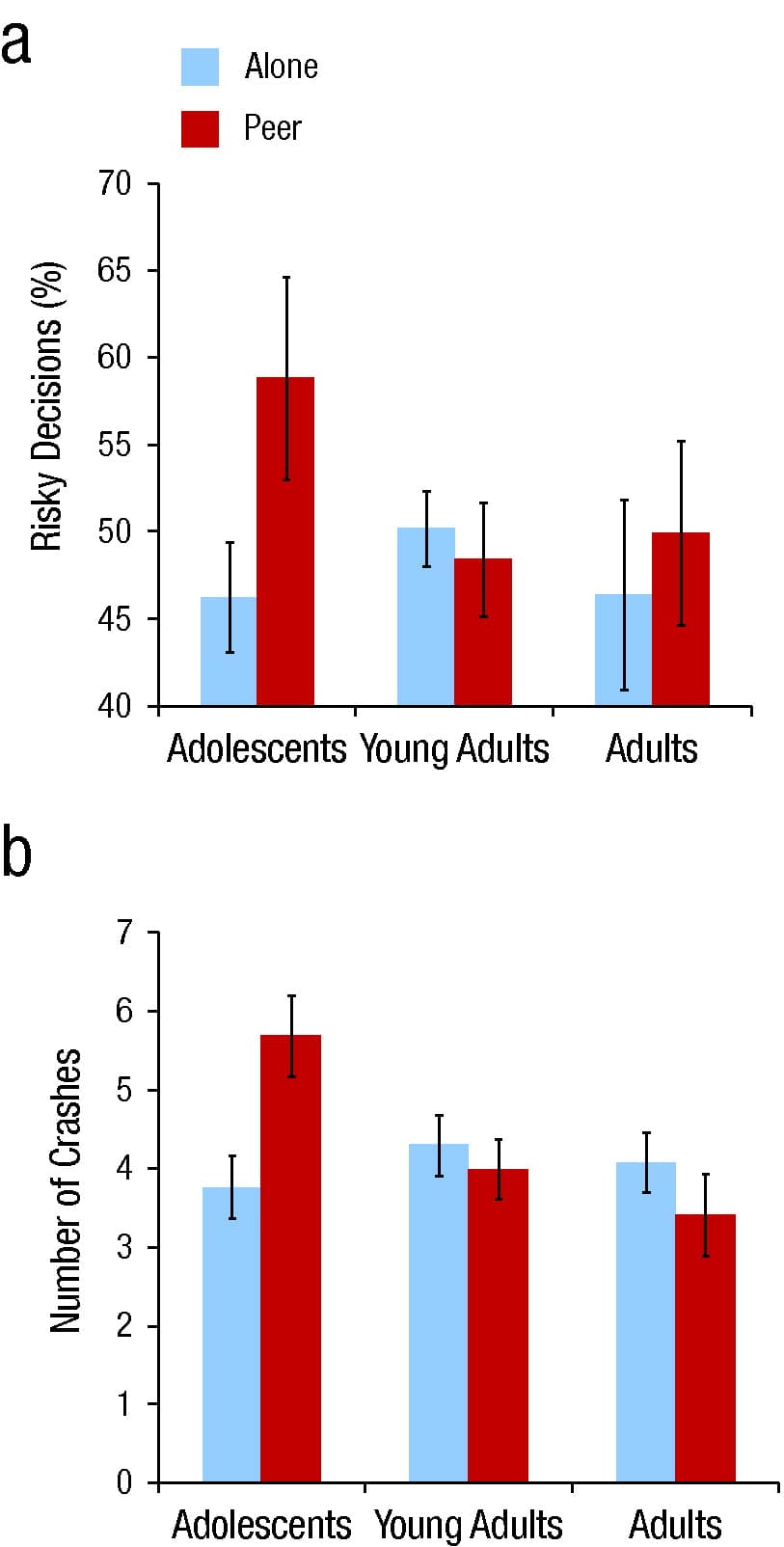
Fig. 2. Susceptibility of adolescents, young adults, and older adults to peer influences on performance in the Stoplight game in Chein, Albert, O’Brien, Uckert, and Steinberg (2011). The graphs show (a) the mean percentage of risky decisions and (b) the number of crashes for participants playing the Stoplight game either alone or with a peer audience. Error bars indicate standard errors of the mean.
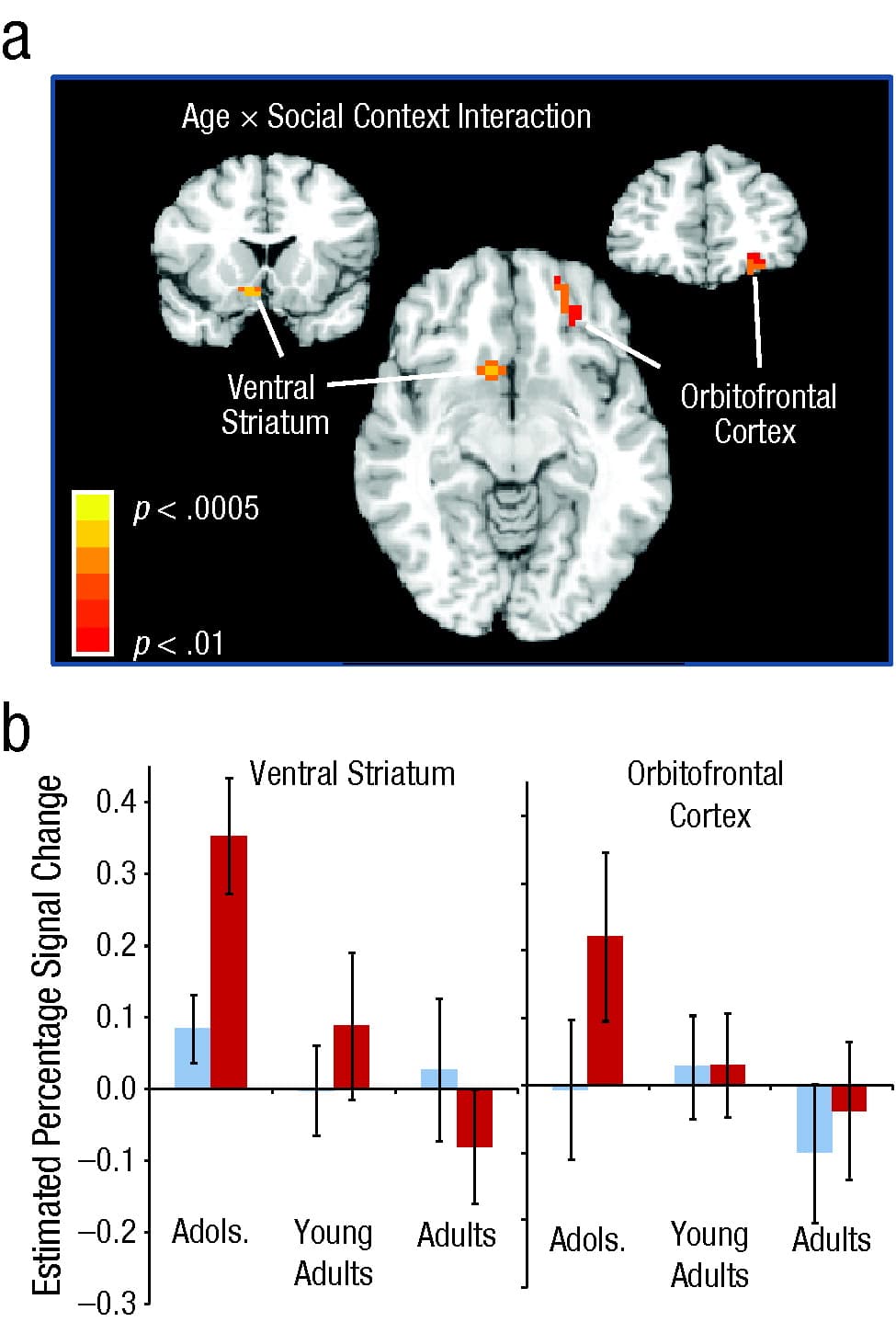
Fig. 3. Brain regions showing an Age × Social Context interaction during the Stoplight game in Chein, Albert, O’Brien, Uckert, and Steinberg (2011). The graphic (a) shows two brain regions exhibiting an Age × Social Context interaction: the right ventral striatum (Montreal Neurological Institute, or MNI, peak coordinates: x = 9, y = 12, z = −8) and the left orbitofrontal cortex (MNI peak coordinates: x = −22, y = 47, z = −10). Brain images are shown by radiological convention (left on right) and thresholded at p < .01 for presentation purposes. The graph (b) shows mean estimated blood-oxygen-level-dependent signal change (standardized coefficients) from the four peak voxels of the ventral striatum and the orbitofrontal cortex in adolescents (adols.), young adults, and adults in alone and peer conditions. Error bars indicate standard errors of the mean.
Indeed, the degree to which both adolescent and adult participants evinced greater ventral-striatum activation in the peer condition compared with the alone condition was inversely correlated with self-reported resistance to peer influence (Fig. 4). These findings constitute the first evidence that peer presence accentuates risky decision making in adolescence by modulating activity in the brain's reward-valuation system.
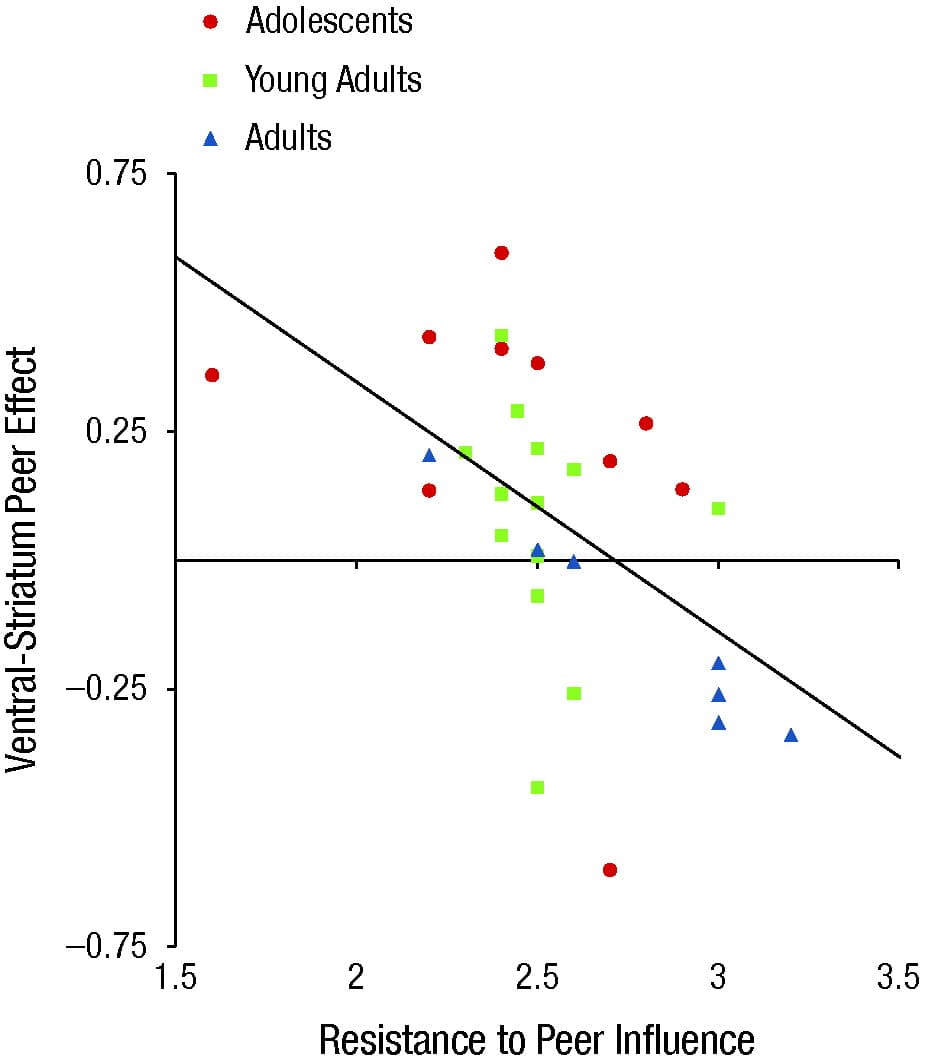
Fig. 4. Resistance to peer influence correlated with right-ventral-striatum activity during the Stoplight game in Chein, Albert, O’Brien, Uckert, and Steinberg (2011). Estimated activity was extracted from an average of the four peak voxels in the ventral-striatum region of interest. The graph shows a scatter plot, with best-fitting regression line, of ventral-striatum activity indicating an inverse linear correlation between self-reported resistance to peer influence and the neural peer effect (i.e., the difference in average ventral-striatum activity in peer relative to alone conditions).
Conclusions and Future Directions
Although our work to date has indicated that the effect of peers on adolescents’ risk taking is mediated by changes in reward processing during adolescence, we recognize that the distinction between risk taking that is attributable to heightened arousal of the brain’s reward system versus that which is due to immaturity of the cognitive-control system is somewhat artificial, given that these brain systems influence each other in a dynamic fashion. Consistent with this notion, in a comparison of children, adolescents, and adults on a task that requires participants to either produce or inhibit a motor response to pictures of calm or happy faces, Somerville, Hare, and Casey (2011) not only found elevated ventral striatal activity for adolescents in response to happy faces, which the authors described as an “appetitive” cue, but also a corresponding increase in failures to inhibit motor responses to the happy (vs. calm) facial stimuli. Thus, adolescents’ exaggerated response to positively valenced social cues was shown to directly undermine their capacity to inhibit approach behavior. Translated to the peer context, this finding suggests that adolescents may not only be particularly sensitive to the rewardsensitizing effects of social stimuli, but that this sensitization may further undermine their capacity to “put the brakes on” impulsive responding.
Despite the promise of this conceptual model, further work is needed to more specifically determine the neurodevelopmental dynamics underlying adolescents’ susceptibility to peer influence and to translate this understanding to the design of effective prevention programs. In an effort to “decompose” the peer effect, we are currently examining age differences in the influence of social cues on neural activity underlying performance on tasks specifically tapping reward processing and response inhibition. In addition, we are investigating whether conditions known to diminish cognitive control (e.g., alcohol intoxication) might exacerbate the influence of peers on risky decision making. Finally, as a first step toward our ultimate goal of using this research to improve the efficacy of risktaking prevention programs, we are examining whether targeted training designed to promote earlier maturation of cognitive-control skills might attenuate the influence of peers on adolescent decision making.