Abstract
The recreational consumption of cannabis has increased significantly across the world with an estimated 180 million people currently using. In the United States, 4.1 million are currently diagnosed with cannabis use disorder. Cannabis dependence and abuse was combined into a single entity as a behavioral disorder with a problematic pattern of cannabis use and termed cannabis use disorder by the Diagnostic and Statistical Manual of Mental Disorders. Chronic use of cannabis has been linked with region-specific effects across the brain mediating reward processing, cognitive control and decision-making that are central to understanding addictive behaviors. This review presents a snapshot of the current literature assessing the effects of chronic cannabis use on human brain function via functional MRI. Studies employing various paradigms and contrasting cognitive activation amongst cannabis users and non-users were incorporated. The effects of trans-del-ta-9-tetrahydrocannabinol (Δ9-THC) in marijuana and other preparations of cannabis are mediated by the endocannabinoid system, which is also briefly introduced. Much variation exists in the current literature regarding the functional changes associated with chronic cannabis use. One possible explanation for this variation is the heterogeneity in study designs, with little implementation of standardized diagnostic criteria when selecting chronic users, distinct time points of participant assessment, differing cognitive paradigms and imaging protocols. As such, there is an urgent requirement for future investigations that further characterize functional changes associated with chronic cannabis use.
Introduction
The recreational consumption of cannabis has increased significantly across the world with an estimated 180 million people currently using. In the United States, 4.1 million are currently diagnosed with cannabis use disorder (CUD). Cannabis dependence and abuse was combined into a single entity and termed as CUD by the Diagnostic and Statistical Manual of Mental Disorders (DSM-5). It is classified as a behavioral disorder with a problematic pattern of cannabis use with clinically significant impairment or distress in a 12 month period with at least two of the following manifestations: (1) cannabis is often taken in larger amounts or over a longer period than was initially intended. (2) There is a persistent desire or unsuccessful effort to cut down or control cannabis use. (3) A significant amount of time is spent in activities necessary to obtain cannabis, use cannabis or recover from its effects. (4) Craving or the urge to use cannabis. (5) Recurrent cannabis use results in failure to fulfill role obligations at work, school or home. (6) Continued cannabis use despite having persistent or recurrent social or interpersonal problems caused or exacerbated by the effects of cannabis. (7) Important social, occupational, or recreational activities are given up or reduced because of cannabis use. (8) Recurrent cannabis use in situations in which it is physically hazardous. (9) Cannabis use continues despite knowledge of having a persistent or recurrent physical or psychological problem that is likely to have been caused or exacerbated by cannabis. (10) Tolerance, as defined by either: (a) a need for markedly increased cannabis to achieve intoxication or desired effect or (b) a markedly diminished effect with continued use of the same amount of the substance. (11) Withdrawal, as manifested by either (a) the characteristic withdrawal syndrome for cannabis or (b) cannabis is taken to relieve or avoid withdrawal symptoms. A subsequent evaluation of adult cannabis users identified nervousness/anxiety, hostility, sleep difficulty and depressed mood among the primary symptoms. Withdrawal signs after abrupt abstinence from cannabis have been quantified previously via a Cannabis Withdrawal Scale reporting that sleep dysregulation, nightmares, and outbursts of anger were the most common or distressful responses. Tests for psychological, sensory and physical symptoms identified irritability and anxiety, cannabis craving, and decreased appetite as withdrawal symptoms that are most pronounced in the first 3 days of abstinence, and strange dreams and difficulty getting to sleep as symptoms that persisted beyond this time frame. Severity of the disorder is graded as either Mild, Moderate, or Severe depending on whether 2–3, 4–5, or 6+ of the above criteria are present. Remission is defined as either (a) early—meeting full criteria previously and not meeting any criteria for CUD for at least three months but less than 12 months (with an exception provided for craving) or (b) sustained—meeting full criteria previously and not meeting any criteria for CUD for at least 12 months or longer (with an exception for craving).
The onset of cannabis use is typically during adolescence with the younger age groups making up the majority of the prevalence and the greatest number of users in the 18–25 year old range. This increase in cannabis use, particularly among youth during stages of substantial brain development has led to growing concerns and controversies arising over its use, its safety, legal status and decriminalization of possession in smaller quantities across many countries. Several studies have reported associations between cannabis use and increased risk for developing schizophrenia-like psychosis. Other reports describe associations between cannabis use and acute/chronic mental health problems, such as anxiety and depression, with severity dependent on frequency, age of onset and genetic vulnerability. Prior evidence also suggests recent smoking and/or blood THC concentrations 2–5 ng ml−1 as being associated with substantial driving impairment, particularly in occasional cannabis smokers. Several neuroimaging studies have yielded important insights into neurobiological changes associated with persistent cannabis use, such as morphological brain changes, differing patterns of brain activity, functional alterations involving cognitive domains and impaired task performance extending beyond the period of intoxication, with earlier onset of use associated with increased alterations in brain structure and function, along with lower IQs.
The current review focuses on functional imaging with respect to hallmarks of drug abuse and dependence to explore the pathology of CUD. Reviews that comprehensively describe addiction circuitries in the brain are available. Our motivated behavior and control is guided by the extraction of salient reward-related information from a large body of stimuli and events. The hedonic characteristics of a specific reward can lead to its exploitation and a sense of pleasure from its consumption, further reinforcing its approach in the future. There have been extensive investigations on the motivation changes that occur for drug and natural rewards in substance use disorders (SUD) along with its neural correlates. The reward threshold of dopamine neurons in the ventral tegmental area (VTA) changes in the initial phases of SUD, resulting in increased firing of these neurons in response to substance-related cues. Studies focusing on cannabis use have shown similar results with cannabis-related cues presented through differing sensory modalities such as auditory-visual, auditory-tactile and visual cues eliciting increases in self-reports of craving in both current and abstinent cannabis smokers. Another avenue of interest to drug abuse is performance monitoring, detecting behavior and making modifications without any reinforcement. This type of cognitive control is vital for our capacity to inhibit reaching for immediate gratification via pleasurable stimuli and adapt our behavior. These processes have been related to deficits in executive control system following chronic cannabis use. The current literature represents a scarcity in studies assessing these cognitive mechanisms exclusively in CUD. Therefore, this review will focus on the overall alterations in brain regions associated with reward processing, cognitive control and decision making after chronic use of cannabis products. We will present a summary of the effects on human brain function as measured via functional MRI (fMRI), incorporating studies employing various paradigms and contrasting cognitive activation amongst chronic cannabis users and non-users.
Recent epidemiological data also indicate increases in treatment demand for cannabis dependence, highlighting the possibility of developing CUD. Current pharmacological and behavioral therapeutic interventions are rather limited though, demonstrating some efficacy. Deficits in daily life functioning during withdrawal appear to be proportional to the degree of cannabis use. Evidence is accumulating on brain function following both brief and prolonged abstinence from cannabis in chronic users, with a specific interest in the aforementioned cognitive mechanisms because of their relevance to treatment compliance and long-term success of interventions. We will also review phase-dependent changes and synthesize the findings at varying stages of abstinence in chronic cannabis users.
The endocannabinoid system (ECS)
The effects of trans-delta-9-tetrahydrocannabinol (Δ-9-THC) in marijuana and other preparations of cannabis are mediated by the endocannabinoid system (ECS), which we will briefly introduce before discussing the fMRI insights into cannabis use. Excellent reviews describe the ECS in addiction processes and brain circuitry. The ECS is comprised of CB1 and CB2 cannabinoid receptors, endogenous cannabinoid agonists arachidonylethanolamide (anandamide) and 2-arachidonoylglycerol (2-AG), referred to as "endocannabinoids," and the enzymes responsible for their production and degradation. Cannabinoid receptor signaling was demonstrated to be G protein-mediated as pertussis toxin-sensitive modulation of cyclic AMP accumulation in rat brain slices. Their identity as 7-transmembrane receptors were later confirmed by cloning of CB1receptors from a rat cerebral cortex by Matsuda and colleagues and CB2 receptors from immune tissue by Munro and colleagues. Cannabinoid receptor activation results in an array of biochemical responses that are dependent on the cell type, such as the inhibition of voltage dependent calcium channels and adenylyl cyclases, resulting in lower cAMP levels and activation of potassium channels, phospholiphases and mitogen-activated protein kinase pathways.
The distribution of cannabinoid receptors is similar in primate and rodent brain, with highest levels in the hippocampal complex, entorhinal and cingulate cortices, frontal gyrus, amygdaloid complex, substantia nigra and cerebellar molecular layer. They are relatively high in associative regions of frontal and limbic cortices, and low-to-moderate in primary and secondary motor and sensory cortices. CB1 receptors have a high abundance particularly in the cortex, basal ganglia, hippocampus, and cerebellum. The majority of CB1 receptors are present on axon terminals and pre-terminal axon segments, while sparing the active zone. Cortical and hippocampal CB1 receptors are localized on cholecystokinin positive GABAergic interneurons (low threshold spiking interneurons) and are expressed in distinct numbers on glutamatergic neurons. They are widely localized in medium spiny neurons in the dorsal and ventral striatum (VS) of the basal ganglia and on axonal pathways entering the globus pallidus heading towards the substantia nigra. In the cerebellum, CB1 receptors are found in parallel and climbing fibers, as well as basket cells. The functionally relevant expression of CB1 receptors have also been identified peripherally in testis, adrenal gland, heart, lung, prostrate, bone marrow, thymus and tonsils. In accordance with these distributions, CB1 receptors modulate feeding, energy expenditure, reward, memory function, perception, movement along with mood-enhancing effects of cannabis or negative, dysphoria-like effects and metabolic homeostasis. CB1 receptors are also found in the thalamic and hypothalamic nuclei, as well as brainstem. CB1 cannabinoid receptors were quantitated by PET scanning methods and found to be diminished in chronic frequent marijuana smokers. The decline in receptor density was pronounced in cortical regions, with the extent of down-regulation correlating with duration of cannabis use. However, after four weeks of monitored abstinence, the CB1 receptor levels returned to that of normal subjects.
Endogenous and exogenous cannabinoids
Endocannabinoids are endogenous lipids produced in the brain that activate cannabinoid receptors. There are also exogenous sources from plants, referred to as phytocannabinoids, the sources of which come from marijuana and synthetic laboratory cannabinoids. The first and broadly studied endocannabinoids are anandamide and 2-AG. Their precursors are present in lipid membranes, and upon activation of certain G-protein coupled receptors or depolarization, endocannabinoids are liberated into the extracellular space. The characterization of the synthesis and degradation pathways for endocannabinoids is starting to yield novel targets for new pharmaceutical agents to manipulate brain endocannabinoid levels, and is beginning to illustrate the role of this system in psychiatric disorders.
Anandamide also activates non-cannabinoid receptors, such as transient receptor potential vanilloid 1 (TRPV1) mediating its vasodilator role and PPAR α and γ receptors with effects on gene transcription. The first phytocannabinoids, i.e. Δ-9-THC, a well-established psychoactive component, and putatively non-psychoactive cannabidiol, were isolated from marijuana by Mechoulam. Of note, marijuana today has important differences compared to marijuana described initially. Initially, the most readily available type of cannabis was resin or hash, followed by traditional herbal cannabis, and then sinsemilla or skunk. Sinsemilla has increasingly taken over the market with rising potency levels of THC concentration and much lower levels cannabidiol, which has implications for studies from previous decades suggesting few cognitive effects of cannabis. In particular, Forti et al found that individuals presenting with first episode of psychosis had smoked higher-potency cannabis and with greater frequency compared to healthy controls.
Functional MRI and Cannabis Use Disorder: Reward processing & cognitive control
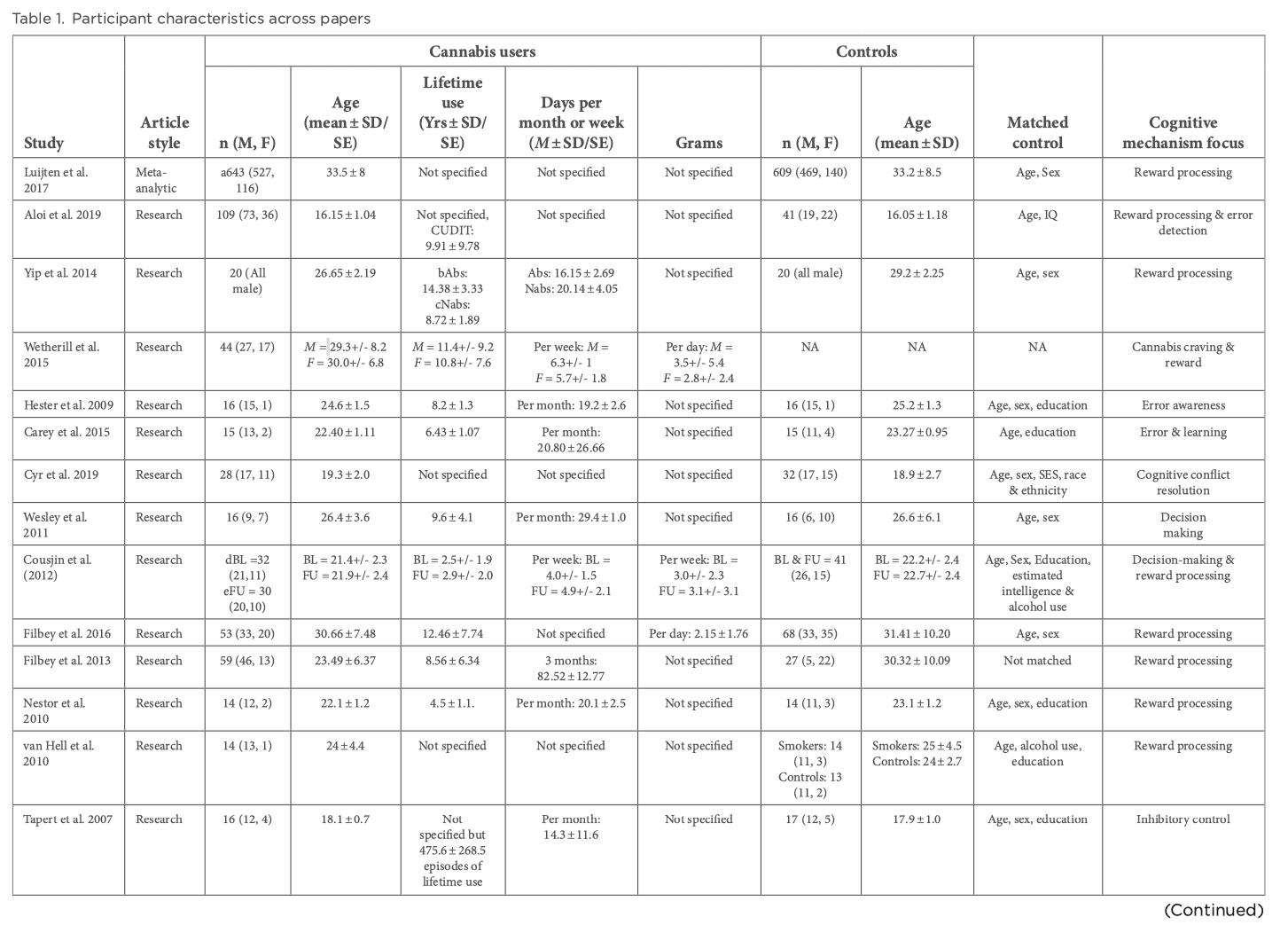
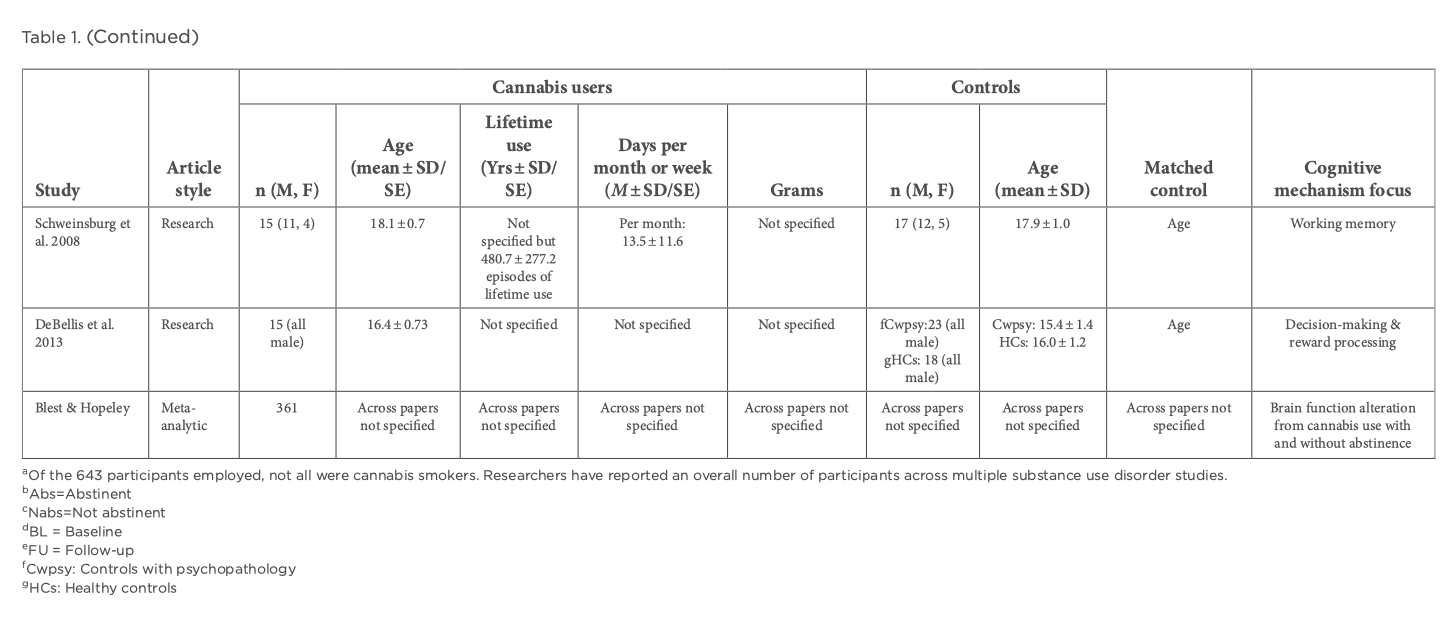
Reward processing entails the anticipation of reward and reward outcome. Reward anticipation arises from the perception of appealing cues with incentive values that have been learned through reinforcement via positive outcomes. In contrast, reward outcomes are pertinent to learning processes and signaling the salience of new stimuli. Reward processing networks include several key brain areas, including the VTA, limbic regions, specifically the VS and frontal areas. CB1 receptors have been located in the VTA, striatum and prefrontal cortex (PFC) and animal models have demonstrated the effect of cannabis on the reward system, with acute exposure to THC and anandamide increasing dopaminergic transmission in the nucleus accumbens. A meta-analysis by Luijten et al aimed to analyze fMRI studies on brain reactivity to nondrug, monetary rewards to understand the nature of reward processing dysfunction in addictive behaviors by separately assessing both reward anticipation and reward outcome. They observed decreased striatal activation during reward anticipation across individuals with SUD compared to healthy controls. During reward outcomes, they found increased activation in the VS across substances. A recent study by Aloi et al used the monetary incentive delay (MID) task to assess reward sensitivity and response accuracy. Study-specific details including demographic information are shown in Table 1. They utilized the CUD identification test (CUDIT) to capture lifetime cannabis use and found increasing scores to be associated with decreased BOLD activation in putamen during feedback on inaccurate trials compared to accurate trials and putamen and anterior cingulate cortex (ACC)/dorsomedial PFC (dmPFC) during feedback on inaccurate punishment trials compared to all trials. CUDIT score was unrelated to striatal modulation by reward. Yip et al found their cannabis-dependent male participants prior abstinence to demonstrate greater VS activation when processing trials where losing had to be avoided. However, only male participants were recruited by this group, therefore potential sex differences in neural responses to cannabis cues were not studied. Only one report by Wetherill et al found differential neural responses to backward-masked cannabis cues versus neutral cues in treatment-seeking, cannabis-dependent adults. They found females to show a positive correlation between neural responses in the bilateral insula and cannabis craving and negative correlation between responses in the left lateral orbitofrontal cortex (OFC) and cannabis craving. Comparatively, males showed a positive correlation between responses in the striatum and cannabis craving. These differential relationships need to be replicated by other groups using non-drug paradigms to understand the overall pattern of reward processing functioning amongst the sexes. The participants investigated in all of these analyses had either psychiatric comorbidities present such as attention-deficit/hyperactivity disorder (ADHD), depression and schizophrenia or a history of exposure to other substances. ADHD, depression and schizophrenia have shown to be associated with alterations in striatal activity during reward anticipation and affect reward processing along with their pharmacological treatment having an impact as well. Although some papers indicate consistent striatal hypoactivations during anticipation in patients without any comorbidities or no associations between CUDIT scores and psychiatric diagnosis, the possibility of these having an effect on reward processing cannot be completely ruled out. Exposure to other substances such as cocaine, opiates, alcohol and nicotine can bring about differing neurobiological changes and so the extent of dysregulation to reward processing networks and consequently to reward anticipation or outcome caused solely by CUD is unclear.
Detecting failure is critical for performance monitoring to learn and adjust to changing situations. Hester and colleagues found their chronic cannabis participants to display poorer awareness of errors during an error detection task compared to controls, this deficit occurred in the absence of a performance deficit in that both groups made similar number of inhibitory control errors. This dysfunction was associated with diminished BOLD activation in ACC, right insula, bilateral inferior parietal and middle frontal regions. These papers are limited by a lack of further follow-up with participants to capture task performance and the nature of learning that may occur in chronic cannabis users after negative feedback. Only one paper was found by Carey et al which examined error processing and learning from errors by utilizing a paired associate learning task. Users showed significantly poorer accuracy in recall along with lower rates on errors corrected on subsequent trials. This reduced error correction rate correlated with diminished BOLD activity in bilateral dorsal ACC, left hippocampus, left putamen, bilateral thalami and bilateral inferior parietal lobules (IPL). Although these findings reveal novel insights towards cannabis users’ sensitivity to negative feedback and consequent learning, the smaller sample sizes employed in these investigations prevents conclusions to be extended further to the rest of the chronic cannabis using population. Both of these investigations did not include a formal diagnostic criteria on abuse and/or dependence and so there is a lack of framework in which the findings can be interpreted because there is no standardization in sample selection and the profile of chronic use cannot be captured.
Withdrawal and abstinence in Cannabis Use Disorder
Compulsion towards drug-seeking after the drug is no longer experienced as pleasurable indicates a potential disturbance in self-regulatory control. This deficit may interfere with a chronic user’s capacity to integrate in rehabilitation programs. Following 12 h, Cyr et al examined the neural correlates of cognitive interference resolution in cannabis-using youth (see Table 1 for study-specific details including demographic characteristics). Their objective was to capture a problematic pattern of cannabis use so they incorporated participants who met at least one DSM-IV substance abuse/dependence criteria with cannabis use at least twice weekly. Seventeen individuals met criteria for abuse, and 12 for cannabis dependence. They used the Simon Spatial Incompatibility task where participants were shown either a leftward or rightward pointing arrow on each trial that matched or did not match with its actual position on the monitor. They had to respond as quickly and accurately as possible to the direction of the arrow using a select finger for left and right directions. The authors found similar performances of cannabis users and healthy controls but users displayed decreased activation in frontostriatal regions. These entailed: bilateral dorsal and ventral PFC regions, ventromedial PFC (vmPFC), middle cingulate, precuneus, parietal lobe, precentral gyrus, left paracentral lobule, pallidum, thalamus, occipital gyri and right putamen in response to conflict. Controls on the other hand showed greater activation in response to incongruent versus congruent stimuli in these regions and cannabis users did not differentially activate these regions to resolve conflict. More activation in the vmPFC was associated with greater number of days of abstinence from cannabis-use prior to scanning. No group differences were detected in the functional connectivity between frontostriatal regions. Wesley et al used the Iowa Gambling task (IGT) to evaluate decision-making. Participants selected random cards from four decks under ambiguous conditions. Each selection produced either a monetary gain or loss. Eventually, based on the wins and losses associated with each deck, two of the decks emerged as resulting in favorable outcomes and two as unfavorable. The unfavorable decks produce larger gains but larger losses too over time while the favorable decks lead to smaller immediate gains but also smaller losses over time. Participants evaluated win and loss contingencies associated with deck choices, and developed decision-making strategies in later phases of the task. Chronic cannabis users performed poorly, failing to develop decision making strategies that resulted in favorable outcomes. During the evaluation of wins and losses, users had smaller BOLD activity in the ACC, vmPFC and parts of superior medial frontal cortex. During the evaluation of losses, users had less activity in these regions as well as the precuneus, posterior cingulate cortex (PCC), superior parietal lobule (SPL) and portions of the dorsal cerebellum and occipital cortex compared to controls. BOLD activity during loss evaluation in ACC, vmPFC and rostral PFC correlated with improved performance in controls but not users. The authors interpreted these findings in the context of failure of cannabis users to develop successful decision-making strategies due to insensitivity to early monetary losses. These findings however are limited by the presence of pre-existing psychiatric comorbidities present in their sample.
After 24 h of abstinence, Cousjin et al examined the predictive value of decision making for future cannabis use, problem severity and the neural circuitry underlying this process in chronic cannabis users. They used the IGT and incorporated a 6-month follow-up telephone interview on present drug use and nicotine dependence as well. The authors found no performance differences between cannabis users and non-users at baseline during IGT, this performance also did not predict cannabis use or problem severity. Brain activations during the decision phase did not differ between chronic users and controls. There were some notable differences found in the neurocircuitry where higher activations were observed for win evaluation in right OFC, insula and posterior region of the superior temporal gyrus. Weekly cannabis use was associated with win-related activity in insula, VPFC and caudate. Higher win-related activity in superior frontal gyrus (SFG) and higher activity in frontal and ventral temporal lobe during anticipation of disadvantageous decisions was associated with cannabis use increases after 6 months. While this type of follow-up study design assessing the mechanisms that lead to relapse is cardinal to understand cannabis use patterns in chronic patients, a self-reported, telephone follow-up limits the extent of data that can be gathered from these patients. Future studies need to include in-person follow-up evaluations using fMRI components to correlate activation changes at baseline with not only clinical measures of use but also changes in neurocircuitry.
Following a 72 h abstinence period, Filbey et al investigated whether cannabis use disrupts the mesocorticolimbic reward processes. They compared the neural responses to cannabis cues versus natural reward cues, e.g. fruit cues. Their results indicated greater BOLD activity in cannabis users specific to cannabis cues compared to natural reward cues in distinct regions within the mesocorticolimbic reward pathway (OFC, striatum, anterior cingulate gyrus, VTA and precuneus) rather than a generalized hyper-responsivity to all reward stimuli. Filbey et al also assessed the neural mechanisms underlying negative reinforcement in cannabis users. They examined the incentive processes for both negative and positive stimuli in users during withdrawal and studied the relationship of these mechanisms to withdrawal symptoms. They used the MID task and despite similar behavioral task performance during reward anticipation between users and controls, found a difference between the type of incentive and group. Cannabis users had greater neural responses to positive incentive trials where money could be earned versus negative incentive trials where loss of money had to be avoided. This was different from the nature of neural responses seen in controls where greater responses were observed in negative incentive trials versus positive. There were no statistically significant differences in VS responses amongst the two groups, however users showed greater VS activity during reward anticipation i.e. monetary gain trials.
Nestor et al employed participants at varying stages of abstinence ranging from 12 to 504 h (mean abstinence 108 h ± 39.7) and investigated the neural activity in chronic cannabis users during the MID task. Cannabis users demonstrated increased BOLD activity in the right VS for cues that predicted monetary gains which correlated with reported lifetime cannabis use. Decreased activity was observed in the left insula in response to loss and loss avoidance outcomes in users similar to the diminished insular activity and lack of error awareness seen in Hester and colleagues’ paper.
At 7 days of abstinence, Van Hell et al compared neural responses to MID task in chronic cannabis users and found diminished reward anticipation in the nucleus accumbens and caudate nucleus bilaterally, left putamen, thalamus, inferior frontal gyrus (IFG) and SFG involved in motor processing and voluntary movement, cingulate gyrus, inferior and middle occipital gyrus. Enhanced activity during anticipation in users was found in temporal gyrus, cuneus, parahippocampal gyrus. Activity differences between controls and users were found in the caudate nucleus, putamen, thalamus, frontal regions and parahippocampal gyrus. The results, however, are only confined to the male sample used for this analysis. Sex differences in brain activation following abstinence need to be further assessed.
At 21 days of abstinence, Yip et al found users to have significantly higher VS activity during MID task during losing outcomes and not at reward anticipation. Those who did not undergo abstinence displayed greater activation within the caudate while processing losing outcomes compared to healthy controls and abstinent cannabis dependents.
After 28 days of monitored abstinence, Tapert et al utilized a go/no-go task to assess response inhibition in adolescent cannabis users and non-users. Participants were asked to withhold a response to an infrequently occurring stimulus and users showed increased BOLD activity in bilateral anterior SFG and middle frontal gyri, right side particularly extending into anterior insula, right IFG, medial PFC, bilateral posterior parietal cortex (PPC), right lingual gyrus, right SPL, IPL and medial precuneus during both inhibitory and non-inhibitory trials of go/no-go task. Users did not show decreased activity compared to controls on either trials of the task.
A critical facet of decision-making is our capacity to retrieve the appropriate information we have gathered from our evaluation of consequences and to maintain and manipulate this right in that moment whilst developing a strategy. Schweinsburg et al focused on a spatial working memory task to assess vigilance and working-memory in cannabis-using teens and non-abusing controls following 28 days of monitored abstinence. They observed similar task performance but group differences in neural responses with decreased right dorsolateral PFC (DLPFC), and increased right PPC during the working memory portion of the task, and increased medial superior and inferior occipital cortex (OC) responses during the vigilance portion.
At full remission, De Bellis et al studied the neural mechanisms of decision-making and reward response in adolescent CUD. Diagnoses were made via the participant and legal guardian administration of Schedule for Affective Disorders and Schizophrenia for School Aged Children Present and Lifetime Version. They used the decision-reward uncertainty task which is a monetary reward task assessing three conditions: reward risk, behavioral risk and no risk. Reward risk brings about a certainty on the decision required to earn a reward but the actual reward outcome is unpredictable, relying on probability. In contrast, behavioral risk brings about an uncertainty on the decision required to earn a reward and no risk brings complete certainty with knowledge of the exact decision required to earn a reward. During risky decisions with uncertainty, increased activations were found in left SPL, left lateral OC, bilateral precuneus in the CUD group compared with controls. Further decreased activations were found in the left OFC to reward in abstinent participants but increased activations in left OFC to no-reward outcomes compared to non-using adolescents. There was no comparison of these findings with healthy controls without any psychopathology with no rationale provided behind it. The control group used had psychopathology similar to the CUD group along with recruitment from the same place. However, the authors do not provide profiles or further details on the kind of psychopathology these control had, its etiology or what criteria were used to define similarity in the psychopathology amongst the two groups.
Blest and Hopley et al conducted a meta-analysis to investigate whether altered brain function associated with chronic marijuana use persists after a sustained period of abstinence to the extent that cannabinoid metabolites were no longer detectable in urine. They assessed brain activation differences between abstinent cannabis users, current users and non-users. Persisting functional alterations in various central executive (DLPFC and PPC) and default mode network (inferior parietal cortex and precuneus) components were found in adolescent cannabis users even after cannabis metabolites were no longer detectable. The current users in the study comprised of both adults and adolescents and the abstinent group was comprised primarily of adolescents, which prevents inferences on the adult abstaining cannabis population. Nonetheless, these results add to the concern about the vulnerability to residual effects of cannabis products on the developing adolescent brain.
Taken together, these data suggest possible sensitivity of regions within the mesocorticolimbic pathway in persistent use of cannabis, namely, VS, OFC, cingulate gyrus, VTA and precuneus. Other regions implicated are putamen, insula, middle and superior frontal, PFC, paracentral lobule, precentral gyrus, parietal regions, occipital, caudate nucleus, nucleus accumbens, hippocampus and thalamus. However, much variation exists in the current literature regarding these functional changes with some groups reporting impairments and other groups failing to replicate or extend such findings. The methodological issues inherent in these papers prevents these findings to be interpreted any further in the context of chronic use. Firstly, there was no utilization of a formal diagnostic criteria on abuse and/or dependence during participant selection in the studies led by Cousjin, Nestor, VanHell and Tapert. Thus, there is no standardization amongst these papers of chronic use and no clear distinction between dependence and abuse. Further investigations are warranted focusing on participant recruitment under a standard framework or diagnostic criteria of chronic cannabis abuse and dependence. Various papers had ranges of abstinence present in their currently using cannabis group which clouds results looking into the effects of chronic cannabis use in the absence of withdrawal. Furthermore, accounts of abstinence in most of the papers analysing effects of withdrawal are based on self-reports and the authors have not verified the accuracy of self-reported recent cannabis use via quantification of THC metabolites either via urine toxicology analysis, gas chromatography or mass spectroscopy. In all of the papers, studying both chronic use with and without abstinence, regular cannabis use and lifetime exposure was gathered through self-reporting inventories which may be exposed to underreporting and recall bias. Users may minimize actual use in denial, guilt or suspect rejection from the study upon reports of greater use or use of other psychedelics. More robust measures are required to accurately assess chronic cannabis use. The majority of the abstinence papers also had their participants either using other drugs or did not exclude patients with nicotine use and alcohol use. These prevent the true effects of cannabis use to be distinguished from the effects of other addictive substances. The eligibility criteria for regular amounts of cannabis consumed (days per week, grams per week and lifetime use) differs across each paper and so collective results cannot be drawn from such heterogeneity in dosages amongst patient population. More consistent measures with robust criteria are required to quantify chronic use with respect to these categories. The tasks utilized in these papers along with the imaging parameters vary between studies and may be a contributing factor to the inconsistencies observed. Most of these studies had modest sample sizes which further limit the power of the results and were cross-sectional in nature. There is an immediate need for longitudinal assessments with larger sample sizes to elucidate the trajectory of reward processing, cognitive control and decision-making changes along with their neural correlates across different stages of life, from adolescence through emerging adulthood and beyond. Such investigations would also need to be implemented under abstinence to characterize stage-dependent changes from acute to prolonged phases in order to yield high quality insights on risk factors that contribute to persistent use and return to use.