Abstract
Early life stress (ELS) is defined as an acute or chronic stressor that negatively impacts a child’s development. ELS is associated with substance use and mental health problems. This narrative literature review focuses on sex and gender differences in the effects of ELS on 1) adolescent neuroendocrine development; 2) pubertal brain maturation; and 3) development of internalizing symptoms and subsequent substance use. We posit that ELS may generate larger hormonal dysregulation in females than males during puberty, increasing internalizing symptoms and substance use. Future research should consider sex and gender differences in neuroendocrine developmental processes when studying the link between ELS and negative health outcomes.
1.1. Introduction
Adverse childhood experiences (ACE) (Giano et al., 2021; Leza et al., 2021), early childhood adversity (ECA) (Shonkoff et al., 2021), early life adversity (ELA) (Hoffman et al., 2019), and early life stress (ELS) (Campbell et al., 2022; King et al., 2020) are terms that have been used to describe exposure to life stressors. These stressors typically encompass experiences of abuse, neglect, and conflict (e.g., incarceration, mental illness, substance misuse, domestic violence). Some researchers expand their definition of a “stressor” to include societal and community factors, such as racial or ethnic discrimination, low socioeconomic status (SES), and neighborhood violence. For simplicity within this review, the term ELS is used in place of other acronyms (ACE, ELA, ECA) when discussing prior literature.
Approximately 20% of adolescents in the United States experience ELS (such as physical, emotional, sexual abuse; neglect; interpersonal violence; poverty; hunger; food insecurity; low SES; racial discrimination; or family strife) at some point in their childhood, and these experiences influence health outcomes (Bomysoad & Francis, 2020; Hoffman et al., 2019). A seminal paper by Felitti et al. (1998) began a body of work investigating the connections between negative early life experiences and adult health and wellbeing. The authors found a strong, positive relationship between the number of ELS experienced and adult physical health problems. Adults with a history of ELS display higher rates of poor cardiovascular health, diabetes, high cholesterol, internalizing problems (anxiety, depression), and substance use disorders (SUD) (Bomysoad & Francis, 2020; Clemens et al., 2020; Heim et al., 2008). Though ELS is associated with health outcomes across the lifespan, the present narrative review will focus on its role in sex and gender differences during adolescence (defined by World Health Organization as ages 10–19) in the development of internalizing problems and substance use (Organization, 2001). The term “sex differences” is used within this review to refer to physical and physiological differences, whereas the term “gender differences” refers to identity, social, and behavioral differences. While broadly referred to in binary terms (male/female, man/woman), both sex and gender exist as a fluid mosaic (Joel, 2020; Suen et al., 2020).
The transition from childhood to adulthood is a period of profound brain development, particularly in areas associated with cognition, emotion, and social processes (Shaw et al., 2019; Vijayakumar et al., 2021). During this period, adrenal and gonadal circulating hormone levels increase as the hypothalamic pituitary adrenal (HPA) and gonadal (HPG) axes mature (Marceau et al., 2015; Romeo, 2013; Ruttle et al., 2015). Previous studies have examined how ELS alters HPA and HPG axes in adolescents (Black et al., 2018; King et al., 2020; Phan et al., 2021; Ruttle et al., 2015), but have not probed the relationships between ELS, internalizing problems, and substance use. Existing reviews exploring these relationships (Arain et al., 2013; Chaplin et al., 2018; Heim et al., 2008; Shaw et al., 2019; Sze & Brunton, 2020), often treat sex and gender differences as a side note. To our knowledge, no review has explored the potential neuroendocrine mediators or moderators of these differences in adolescent response to ELS or built a coherent conceptual model to explain the complex relationships and processes by which ELS alters HPA and HPG function, potentially leading to increased internalizing problems and substance use.
This narrative literature review seeks to focus on sex and gender differences in the role of ELS in relation to 1) adolescent neuroendocrine development; 2) neurobiological brain changes accompanying puberty; and 3) development of symptoms of mood disorders and substance use. This review provides a synthesis of recent research from disparate fields of study following an integrative conceptual framework, covering varying combinations of ELS, cross-sectional and longitudinal studies, and research based on animal (Bourke & Neigh, 2011) and human (Ruttle et al., 2015) models. The review concludes with a discussion of the potential buffering effects of resilience factors and implications of this conceptual model for policy, practice, and future research.
2.1. Conceptual Model
As presented in Figure 1, this narrative literature review proposes a conceptual model integrating ELS, HPA and HPG axis function, depression and anxiety symptoms, and substance use in adolescents (Figure 1). In this model, ELS generates chronic stress, leading to HPA axis dysfunction, increasing baseline (not stress-induced) cortisol levels and downregulating negative feedback mechanisms. The consistently high baseline cortisol levels then reduce the capacity of the system to increase signaling in response to an acute stressor. Thus, an abnormally high level of baseline cortisol signaling leads to a hypoactive acute stress response. Sex hormones from the HPG axis increase (estrogens) or decrease (testosterone) baseline cortisol levels (Oyola & Handa, 2017; Sze & Brunton, 2020) affecting the maturation and function of emotion regulation and reward circuitry (Eck & Bangasser, 2020; Farooqi et al., 2018; Frere et al., 2020; Hanson et al., 2021). Additionally, high basal levels of cortisol may contribute to early puberty, which is associated with increased internalizing symptoms in adolescents (Barendse et al., 2022; Natsuaki et al., 2009), who may turn to substance use to cope with stress and other negative emotions. This effect is moderated by sex and gender, as females/girls are more likely than males/boys to display a hypoactive acute stress response after ELS (Bouma et al., 2011; Bourke & Neigh, 2011), decreased HPA and HPG hormone associations after ELS (Black et al., 2018; Ruttle et al., 2015), increased depression and anxiety symptoms in response to stress (Natsuaki et al., 2009), and use of illicit and legal substances to cope with negative mental health symptoms (Kuhn, 2015; Romano et al., 2021). The subsequent sections of this review follow the conceptual model and provide empirical support for the proposed relationships between ELS, affect and substance use, as well as their mediating and moderating factors.
Fig.1
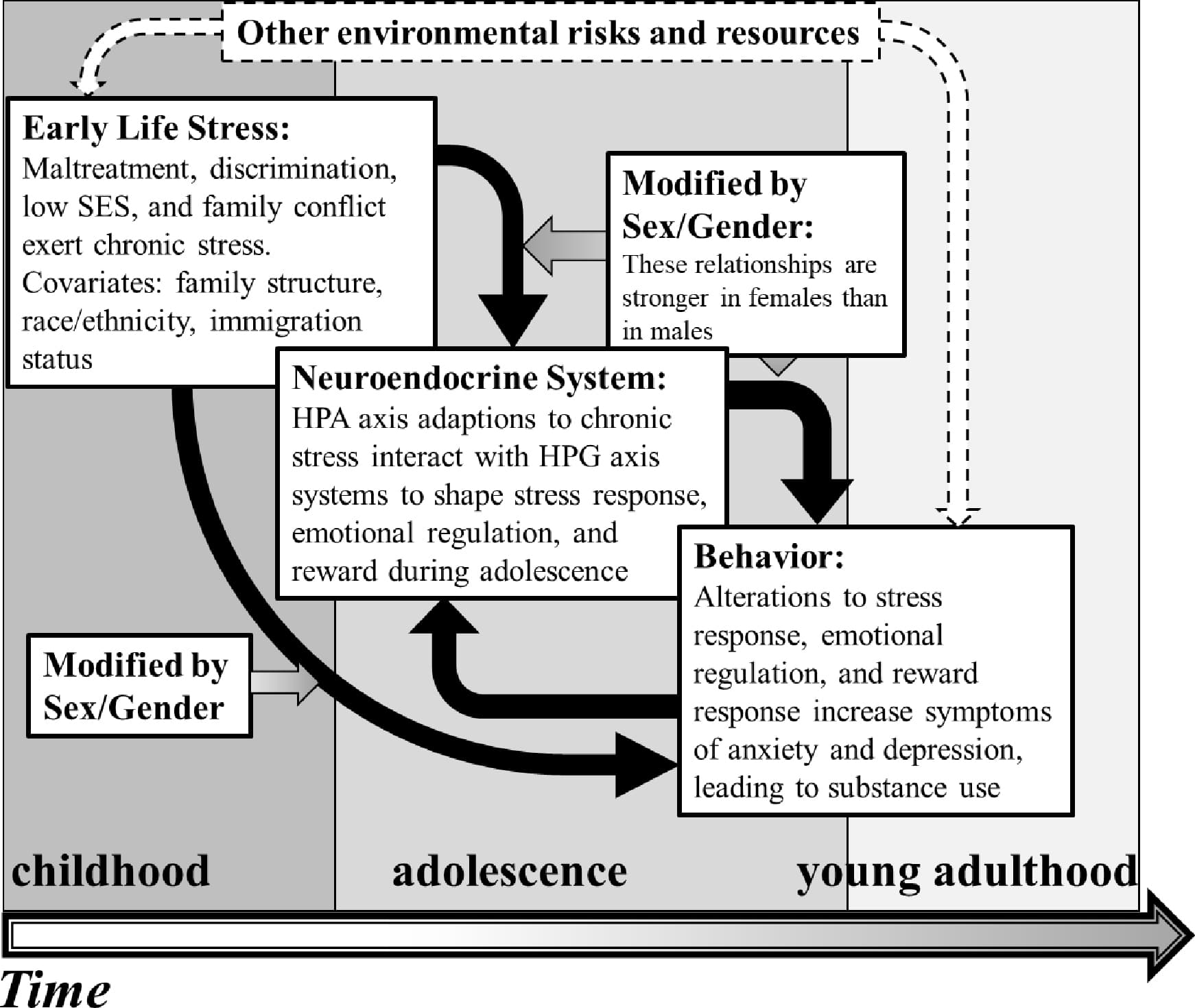
2.2. Early Life Stress: A Starting Point
This review defines ELS as experiences of abuse (physical, emotional, sexual), neglect (physical, emotional), problematic family history (mental illness, substance use, parental separation, family member incarceration), and/or other adverse environmental factors (low SES, neighborhood violence, and discrimination), associated with child development (Assari, 2020; Bomysoad & Francis, 2020; Danyluck et al., 2021; Kuzminskaite et al., 2021; Lee & Santiago, 2021; Murphy et al., 2022; Shonkoff et al., 2021; Turner et al., 2022; Vaughn et al., 2016). These experiences have shown gender differences in prevalence of abuse occurring prior to age 18 (Meng & D’Arcy, 2016; Merrick et al., 2018; Swedo, 2023), with a greater proportion of men reporting physical abuse whereas women are more likely to report sexual abuse. Additionally, a greater proportion of women than men reported growing up in a household with mental illness, substance use, and intimate partner violence (Meng & D’Arcy, 2016; Merrick et al., 2018; Swedo, 2023). The percentages reported depend on the study design and sample; Merrick et al. (2018) report 7% of men and 16% of women in their sample experienced sexual abuse, while Meng and D’Arcy (2016) reported rates of 31% and 69%, respectively. Despite these variations, the trends in gender differences remain the same across multiple studies.
Other analyses of ELS have attempted to identify associations between health outcomes and stressors by stressor type (Giano et al., 2021; Sumner et al., 2019). To this end, Orendain et al. (2023) analyzed variables associated with ELS using data from the Adolescent Brain Cognitive Development (ABCD) study (Volkow et al., 2018). Physical and sexual violence, scarcity, and parental psychopathology were more strongly associated with internalizing problems in youth, whereas neighborhood threat, household dysfunction, and prenatal substance exposure were strongly associated with externalizing problems. However, the overall pattern showed parental psychopathology, household dysfunction, and neighborhood threat to have the strongest correlation with problematic youth behavior (Orendain et al., 2023).
Connections among multiple sources of ELS are expected, as one source of stress often impacts another. For example, maternal depression is associated with more negative parenting behaviors, increased family discord, and parental display of maladaptive coping behaviors (Oldehinkel & Bouma, 2011). Adolescent food insecurity is associated with familial discord, peer bullying, anxiety and depressed mood in both child and parents, child behavioral problems, low self-esteem, and low self-efficacy (Godrich et al., 2019; Leung et al., 2020; Melchior et al., 2009). Finally, growing up in a family affected by mental health problems or substance misuse is associated with a 5–7 fold increase in the odds of experiencing any type of child abuse or neglect (Clemens et al., 2020). Each of these interrelated stressors may vary in type, timing, duration, severity, and relationship of perpetrator to youth, all of which differentially impact mental and physical health (Cohodes et al., 2021). However, such granular information is unavailable in large longitudinal studies (such as the ABCD). Readers interested in analysis of stressor type, timing, etc., are encouraged to read the review of Cohodes et al. (2021). Since the current review utilizes a breadth of research to build an integrative conceptual model, studies cited here will be a mix of both individual and cumulative sources of ELS.
2.3. HPA and HPG Function During Adolescence: Effects of Stress and Sex
Adolescence is a time of physical, emotional, and mental development, during which a person transitions from childhood to adulthood. Essential to this time period is the onset of puberty, the physical process of sexual maturation that is initiated through the HPG axis (Oldehinkel et al., 2011). Activation of the HPG axis results in the gonads releasing estradiol, progesterone, and testosterone into the bloodstream where they act on sex hormone receptors located throughout the body (Acevedo-Rodriguez et al., 2018). The HPG axis is tightly controlled via feedback mechanisms, where estrogen and androgen receptors (ER and AR) in the hypothalamus allow for either positive or negative feedback onto the HPG axis, thereby regulating circulating gonadal hormone levels (Acevedo-Rodriguez et al., 2018; Marceau et al., 2015).
Investigations into the effects of ELS on HPG function tend to center around questions of pubertal timing and tempo (Assari et al., 2020; Herting et al., 2020) or the rise in affective and behavioral problems that occur with puberty (Booth et al., 2003; Dorn et al., 2009). Earlier pubertal onset is associated with poorer mental health outcomes among girls with a history of ELS but not among boys with a history of ELS, suggesting the expression of sex hormones by the HPG axis influences response to stress (Marceau et al., 2015). However, earlier-developing girls also tend to seek out older peer groups, potentially exposing themselves to higher-risk situations that contribute to social stress and poorer mental health outcomes (Mendle et al., 2007).
During adolescence, both the HPA and HPG axes increase their basal hormonal output (Marceau et al., 2015). The HPA axis mediates response to stressors through activating physiologic, immune, and metabolic systems. When a stressor is perceived, neuronal stimulation of the hypothalamus begins a cascade of events that result in the release of cortisol and dehydroepiandrosterone (DHEA) from the adrenal gland into the bloodstream (Herman, 2022; Marceau et al., 2015). DHEA is sulfonated in the brain and liver to make the more abundant stable form, DHEAS (Maninger et al., 2009). Cortisol binds to glucocorticoid receptors (GR) present in many cells throughout the body, and DHEA(S) interacts with multiple receptor types, although a clear DHEA-specific receptor has not been identified (Maninger et al., 2009). For this reason, DHEA is hypothesized to act through conversion to sex steroids, binding both to ER and AR (Maninger et al., 2009). The multiple downstream effects of cortisol and DHEA help the body adapt and respond to a variety of stressors. The response is eventually terminated through a negative feedback mechanism dependent on GR and other receptors in the hypothalamus, hippocampus, and pituitary (Herman, 2022; Sze & Brunton, 2020).
Chronically high levels of HPA axis activity could be associated with a persistent, high baseline measure of cortisol that fails to decrease over the course of the day, as is typical with normal diurnal variation (Murphy et al., 2022). With chronic activation, the HPA axis can no longer mount an adequate response to an acute stressor (Danese & McEwen, 2012). This homeostatic modification increases the “allostatic load” to potentially toxic levels (McEwen & Akil, 2020), generating neuronal damage (Berens et al., 2017; Marceau et al., 2015), an increase in inflammation, and the early pubertal maturation and accelerated biological aging referred to as “stress acceleration” (Berens et al., 2017; Sumner et al., 2019). Sex differences in this pattern of high baseline activity and blunted response to acute stressors are observed across preclinical and human data. For example, adolescent female rats exposed to chronic stress also exhibit blunted corticosterone (rat analogue of cortisol) response to an acute stressor, an effect that persisted into adulthood (Bourke & Neigh, 2011). In a human study comparing adolescent males and females with and without parental depressive problems (a type of ELS), only females with parental depressive problems exhibited a blunted cortisol response to the acute social stressor (Bouma et al., 2011). This suggests ELS has a stronger association with female acute stressor response than male response.
HPA and HPG axes are traditionally thought to exhibit a negative interaction with one another, where the product of one has an inhibitory effect on the other (Son et al., 2022). However, during adolescence the HPA and HPG axes can have a positive interaction with one another, and the coordinated action of both axes contributes to a greater sense of well-being (Chafkin et al., 2022). Cortisol is positively associated with both DHEA and testosterone in children as young as 9 years old (Black et al., 2018; King et al., 2020). This relationship was shown to be less positive among adolescents who experienced high levels of ELS, an effect that was more pronounced in females than males (Black et al., 2018; King et al., 2020). In one prospective longitudinal study, multi-level modeling assessing the relationship between ELS and hormone coordination found high ELS between the ages of 3 and 6 years interacted with sex to predict looser cortisol-testosterone association at age 9 in females but not males (Black et al., 2018). This is consistent with an earlier study by Ruttle et al. (2015) that found decreased association of cortisol and testosterone among females with a history of ELS, but not those without a history of ELS or any males.
Conversely, increases in association have also been observed. For example, a longitudinal twin study found tighter association of DHEA-testosterone in adolescent females than adolescent males. Additionally, the existence of current family psychopathology enhanced the association of DHEA-cortisol in females only (Phan et al., 2021). The study does note that the existence of family psychopathology may not be a strong enough stressor to generate predicted effects on hormone associations. Further, hormones were measured at a single point during the day, rather than examining multiple time points throughout the day, potentially missing any associations not present at the baseline measure. Finally, effects of shared environmental factors were stronger in females than males, accounting for 51% of DHEA variance in females, but only 27% in males (Phan et al., 2021). Thus, family psychopathology (ELS) appears to alter hormone associations more strongly in females compared to males. Females tend to begin puberty before males, and this difference prompted the studies above to incorporate pubertal status into their analysis. The results varied, with some studies reporting puberty was not a predictor of hormone coordination over time (Black et al., 2019; Ruttle et al., 2015), some finding pubertal status to be associated with the strength of the hormonal associations (King et al., 2020), and others finding it to be associated with the direction of the association (Phan et al., 2021). These effects were separate from those of ELS, suggesting that ELS is associated with a non-normative shift in these related hormones.
2.4. Translating from Hormones to Behavior: Development of Depression and Anxiety
2.4.1. Hormone activity in the maturing adolescent brain
Complex processes underlie the relationship between ELS-induced changes in HPA and HPG axis function and increases in internalizing symptoms. One potential target is the maturing corticolimbic system in the adolescent brain. Maturation requires activity-directed synaptic remodeling of neuronal ensembles (Arain et al., 2013), where commonly used connections are strengthened and unused connections removed (Juraska et al., 2013). In addition to this remodeling, myelination increases signal transmission across the cortex and subcortical regions (Shaw et al., 2019). This increased malleability marks adolescence as a sensitive period for maturation of brain and hormonal circuitry, allowing childhood and adolescent experiences to alter hormonal function (Sisk & Gee, 2022). This, in turn, informs the maturation process and leads to modifications of neuronal ensembles that persist into adulthood (Koss & Gunnar, 2018; Murphy et al., 2022; Shaw et al., 2019). Thus, the plasticity required to mature neuronal circuits also leaves them susceptible to insult. For example, cortisol can act on emotional processing centers via GR activity. Expression of GR mRNA increases within the prefrontal cortex (PFC) from childhood to adulthood (Perlman et al., 2007), increasing the population of GR present in developing PFC and enhancing responsivity to cortisol signaling via GR activity. In situations of high cortisol signaling, the excessive stimulation from a hyperactive HPA axis can lead to neurotoxicity, oxidative damage, and neuronal atrophy causing damage to brain regions involved in cortisol signaling (McEwen & Akil, 2020).
Sex hormones act within the brain, organizing and activating connections (Arain et al., 2013; Beck et al., 2023). The HPG axis and its hormones participate in transcriptional regulation of brain plasticity and dopaminergic neurotransmission vital to reward, affect, and motivational neurocircuitry (Eck & Bangasser, 2020). These hormones also aid in developing the immature inhibitory GABA circuits within the adolescent brain that are required for behavioral inhibition and other cognitive functions (Arain et al., 2013; Fruzzetti & Fidecicchi, 2020). Estrogens have myriad abilities to alter neurobiology through interactions with other neurotransmitters such as serotonin, dopamine, or acetylcholine, or directly via genomic activity (McEwen & Milner, 2017). The hippocampus, amygdala, and PFC are sexually dimorphic, responsive to sex hormones, and are a part of the greater stress response circuitry within the brain (Goldstein et al., 2010), undergoing significant development during adolescence (Blakemore et al., 2010). Thus, HPA and HPG axis activity influences adolescent brain development, and dysregulation in either axis can interfere with the maturation of emotional processing and control centers, contributing to the onset of symptoms of depression or anxiety.
2.4.2. Sex and gender differences in depression and anxiety
Prior to puberty, the prevalence of anxiety and depression is roughly equal between the genders (Spielberg et al., 2019). As adolescence and puberty begin, sex and gender differences start to emerge (Altemus et al., 2014). An 18-month study of adolescent males and females ages 9–14 found females exhibited positive correlations between baseline cortisol levels and anxiety. Three cortisol measures were taken every 6 months and the study concluded with an assessment of social and general anxiety. For females, concurrent baseline cortisol levels were positively correlated with symptoms of social and general anxiety, and a greater longitudinal increase in cortisol predicted higher general and social anxiety in females only (Schiefelbein & Susman, 2006). In a cross-sectional study of pre-adolescent and adolescent females aged 8–14, investigators collected data using self-report and clinical interview and found that concentrations of DHEA had a low positive correlation with anxiety symptoms even after controlling for pubertal development (Mulligan et al., 2020), though the researchers note future studies should include adolescent males for comparison. Another cross-sectional study by Natsuaki et al. (2009) found a positive association between heightened cortisol acute response to an interpersonal stress task and higher anxiety and depressive symptoms for females age 11–16 but not males. However, a longitudinal study of adolescents (average initial age=14) found males with elevated baseline cortisol were more likely to develop clinical depression by age 17 than males with low baseline cortisol, an effect that was not observed among the females in the study (Owens et al., 2014). These seemingly contradictory results in the literature come from studies utilizing a variety of measures to assess HPA and HPG axis function, differing populations of at-risk or community samples, and different study designs. In addition, earlier pubertal timing is associated with increased mood and behavioral disorder symptoms (Hirtz et al., 2022; McNeilly et al., 2022; Natsuaki et al., 2009; Oldehinkel et al., 2011), which may contribute to the variation in results.
As adolescents progress through puberty, changes in their developing body can become a source of stress. Increases in self-consciousness and anxiety over peer evaluation and acceptance can generate negative feelings surrounding physical appearance, an effect observed to a greater extent among girls than boys (Oldehinkel & Bouma, 2011; Oldehinkel et al., 2011), leading to increases in rumination and symptoms of depression. Grabe et al. (2007) found that both adolescent girls and boys engaged in self-objectification, with a desire to conform to Western body ideals, yet only girls showed a significant association with increased rumination and depressive symptoms two years later. Gender roles encouraging boys to gain agency through athletic and academic goals may protect boys from the perceived lack of control over body and environment experienced by girls (Grabe et al., 2007). Thus, the burden of new societal expectations based on a more mature physical appearance may generate increased depression and anxiety symptoms to a greater extent among girls than boys (Oldehinkel et al., 2011). This may contribute to the gender difference in prevalence seen in adulthood, where women are twice as likely as men to have depression or anxiety (Eck & Bangasser, 2020; Oldehinkel & Bouma, 2011; Spielberg et al., 2019).
2.4.3. Impact of ELS
ELS is associated with increased prevalence of adult anxiety and depression, especially among those who experienced higher numbers of ELS (Danese & McEwen, 2012; Heim et al., 2008; Kuzminskaite et al., 2021). Studies have shown that this increased prevalence appears as early as adolescence. Adolescents show a positive association between the level of ELS experienced (number of stressors) and the odds of having a current mental health diagnosis (Bomysoad & Francis, 2020). When compared to adolescents exposed to no stressors, exposure to one stressor increased the likelihood of a depression diagnosis by almost 2.5 times and exposure to 4+ stressors increased the chances of a diagnosis by 10 times (Bomysoad & Francis, 2020).
ELS may exert additional negative effects on adolescent girls as compared to boys. A longitudinal study of adolescents comparing across gender and ELS found girls in the highest category of ELS maintained the highest levels of depressive symptoms at ages 14 and 17. Boys in the highest category of ELS exhibited high depressive symptoms at age 14, but not at age 17 (St Clair et al., 2015). In a longitudinal study of adolescent males and females, ELS predicted decreased functional connectivity between the hippocampus and PFC for both sexes, and between the amygdala and PFC for females only. The authors noted this “double hit” may partially account for the higher levels of internalizing symptoms among girls compared to boys within their study (Herringa et al., 2013).
Overall, these studies suggest sex and gender differences in the magnitude of association ELS has with internalizing symptoms, potentially via hormonal inputs during the process of maturation of the corticolimbic system. These alterations likely persist through adulthood, accounting for the increased rates of internalizing disorders among persons with a history of ELS (Clemens et al., 2020; Heim et al., 2008).
2.5. Using Substances to Cope: From Experimentation to Problem Use
2.5.1. Adolescent substance use
Adolescence is normatively associated with an increase in substance experimentation and use; a cross-sectional study of a nationally representative sample found 20% of 12th grade students report recent (past 30 days) use of any illicit substance compared to 6% of 8th grade students (Johnston et al., 2022). Before use is initiated, attitudes toward substance use can serve as indicators of risk. A cross-sectional study of early adolescents found perceived peer use to be the strongest indicator of curiosity to use alcohol and nicotine (Martz et al., 2022), noting that adolescents tended to overestimate the number of peers using substances.
The perception of peers using substances combined with the intensity of social pressures and influence experienced during adolescence may lead to substance experimentation (Martz et al., 2022). Developmentally, the adolescent brain is primed to seek out novel experiences and take risks (Compton et al., 2022), and exhibits an enhanced susceptibility to reward. Multiple animal studies have found the rewarding properties of alcohol, nicotine, and psychostimulants to be increased in adolescent vs. adult animals (Kuhn, 2015). This heightened response to reward and adolescent propensity toward risky behaviors are likely driven by the immature circuitry present in the PFC (Compton et al., 2022). The adolescent PFC exerts insufficient inhibitory power over a highly active reward center (Peper et al., 2013; Welker et al., 2015).
Reward circuitry is also sensitive to HPA and HPG axis activity. Stress and sex hormone receptors are present in reward circuits, which suggests sensitivity to ELS (Hanson et al., 2021; Welker et al., 2015). Substance use is associated with higher baseline cortisol levels in adolescents, an effect moderated by stressful life events (Rao et al., 2009). Among adolescents with high ELS, use of alcohol by age 14 was associated with high baseline cortisol and blunted acute cortisol response to a social stressor (Rahal et al., 2022). Further, ELS alteration of HPG axis function leads to a dose-dependent increase in the likelihood of early menarche (Henrichs et al., 2014), which in turn is associated with earlier initiation of substance use (Downing & Bellis, 2009; Kim et al., 2017; Kuhn, 2015; Stice et al., 2001).
2.5.2. Coping with negative emotions
What begins as substance use experimentation can become a coping mechanism for ELS, especially in the absence of positive coping role models and learned coping behaviors (Marceau & Abel, 2018; Molina & Whittaker, 2022; Wadsworth, 2015). Experimental initial use can evolve into a desire to suppress or escape negative emotions (Belisle & Dixon, 2020; Gerrard et al., 2018; Heitzeg et al., 2018), such as experiences of perceived discrimination, which are positively associated with using substances to cope among Black and LGBTQ adolescents (Amaro et al., 2021). This pattern increases the frequency of substance use, as persistent stress leads to a continued need to cope (Grummitt et al., 2021; Heitzeg et al., 2018; Martz et al., 2022). The cycle is evident in a longitudinal study of adolescent e-cigarette and traditional cigarette users, which found that higher depression and anxiety symptoms were associated with a 25% (e-cigarette) and 19% (combustible cigarette) increase in the odds of continued nicotine use one year later. In addition, continued cigarette use increased the odds of anxiety and depressive symptoms by 7% two years later among both combustible and e-cigarette users (Lee et al., 2023). The authors of this study suggest that youths were using nicotine as a coping mechanism and that prolonged use prevented adoption of other positive coping mechanisms, though the negative effects of nicotine on stress response systems were also noted as a potential factor for increased odds of depressive and anxiety symptoms with extended use (Lee et al., 2023).
Substance use is more widely associated with internalizing symptoms among adolescent girls than among boys, suggesting a gender difference in motivation for use. Girls are more likely than boys to use alcohol to cope rather than to “have fun” (Agoglia et al., 2020; Romano et al., 2021) and are more likely than boys to use cigarettes to relieve anxiety (Kuhn, 2015). For example, a cross-sectional study of adolescent boys and girls showed positive associations between severity of depressive symptoms and frequency of consumption and increased intoxication in both genders, but this association was stronger in girls (Johannessen et al., 2017). Boys with the highest depressive symptoms had 1.6 increased odds of frequent intoxication, whereas girls in the same category had 2.1 increased odds. Girls also showed a positive association between anxiety symptom severity and measures of alcohol consumption and intoxication that was not present in boys (Johannessen et al., 2017). This is in contrast to historical measures of adolescent substance use in which boys are more likely to use than girls (Wade et al., 2022). Conversely, a cross-sectional study of adolescent boys and girls revealed that girls with greater self-reports of mental well-being were significantly less likely to engage in substance use to cope with COVID-19 related changes -- such as isolation or stress -- an effect not observed among the boys in the study (Romano et al., 2021). Taken together, these studies point to a greater association between mental health and substance use among adolescent girls. Notably, the above studies do not focus on the role of ELS in the development of symptomology leading to substance use, and adaption of substance use as a coping response can vary by substance and race (Assari et al., 2018).
A history of ELS is associated with earlier initiation of substance use (Bardo et al., 2021; Compton et al., 2022). There is an inverse relationship between number of stressors experienced and age of substance use initiation (Rothman et al., 2008), suggesting early adoption of substances as a coping mechanism for ELS. Indeed, those with a higher number of ELS were more likely to report initiating drinking to cope rather than for social and pleasurable reasons, and at an earlier age than those with fewer ELS (Rothman et al., 2008). Adults who engage in heavy polysubstance and opioid use report earlier ages of initiation, greater use of drugs to avoid negative emotions, and higher rates of ELS than less frequent users (Sadeh et al., 2021).
Females have been observed to progress more rapidly from experimentation to heavy substance use (Eck & Bangasser, 2020; Georgieva et al., 2022; Heitzeg et al., 2018; Kuhn, 2015), and girls who begin drinking alcohol in early adolescence (ages 11–14) escalate to heavy alcohol use faster than same-age boys (Cheng & Anthony, 2018). Among a sample of adults with opioid use disorder, 82% had experienced some type of ELS, and of those with 3+ stressors, significantly more women than men had a comorbid mental health condition (Evans et al., 2020), illustrating a stronger association between ELS and adult mental health outcomes. Additionally, a study of emergency department patients found a stronger association between the number of ELS and substance use among women compared to men (Cunradi et al., 2020). Finally, in a nation-wide sample, the predicted probability of a drug use disorder among adult women with a history of 3 + ELS was significantly higher than among adult men with a history of 3+ ELS (Evans et al., 2017).
Cumulatively, these studies indicate that ELS is associated with higher risk of using substances to cope with depression and anxiety symptoms in adolescence, and that it does so to a greater extent for girls than for boys. As illustrated in Figure 1, the proposed pathway behind this sex and gender difference is as follows: ELS is associated with greater alterations of HPA and HPG axis function in adolescent females, which echoes into the maturation of the corticolimbic structures of the brain, increasing the existing propensity for depression and anxiety, which may drive girls with a history of ELS to use substances as a coping mechanism. This highlights that it is critical to intervene during adolescence, a sensitive period that may allow for a recovery of hormonal and neurodevelopmental function (DePasquale et al., 2021).
2.6. Other Environmental Risks and Resources
2.6.1. Peer influences
Some of the connection between ELS and internalizing symptoms (and subsequent substance use) may be due to peer influences. Early pubertal maturation and early maturing adolescents are more likely to seek affiliation with an older peer group; these affiliations may expose such youth to situations they are not cognitively prepared to handle, despite the physical indications of maturity (Downing & Bellis, 2009). Accordingly, association with deviant peers is a potent predictor of substance use initiation (Kuhn, 2015; Vijayakumar et al., 2023). Studies have reported peer use of substances to be a risk factor for girls, for boys, and for both (Amaro et al., 2001; Grummitt et al., 2021; Martz et al., 2022). Additionally, bidirectional associations between substance use and endocrine function (indicated in Figure 1) may exacerbate stress response to adolescent social conflicts. Substance use alters HPA and HPG axis baseline output and responsivity to acute stressors (Marceau & Abel, 2018; Milivojevic & Sinha, 2018; Tweed et al., 2012; Vuong et al., 2010), and continued use of substances to cope with these stressors may increase risk of developing internalizing symptoms. Thus, peers and peer conflict are significant factors in the development of substance use in adolescence.
2.6.2. Genetic vulnerability
Genetic and epigenetic pathways by which ELS increase the risk of anxiety, depression, and substance use are represented in our model by the arrow directly connecting ELS to these behavioral outcomes. For example, polymorphism of the serotonin transporter gene 5HTTLPR moderates the relationship between depression and anxiety symptoms and ELS, with the short allele conferring risk and the long allele resilience (Heim et al., 2008; Lewis et al., 2016; Schiele & Domschke, 2018). Substance use and progression to SUD has also shown heritability, with studies identifying an “addiction gene set” (Hatoum et al., 2023; Marceau & Abel, 2018). Many genes within this set are also part of the cortisol regulation pathway, implicating involvement of the HPA axis in SUD. Additionally, there was an enrichment of genes involved in estrogens signaling, suggesting both the HPA and HPG axes contribute to the genetic vulnerability to substance use.
2.6.3. Intergenerational transmission of ELS
The effects of ELS can span multiple generations. Mothers with a history of ELS are more likely to report food insecurity, low SES, and interpersonal violence (Chilton et al., 2017). The children of mothers who experienced ELS are more likely to experience ELS themselves than children born to mothers without a history of ELS (Smith et al., 2021). A study of parent-child dyads showed an increase in likelihood of child arrest (1.78 times) and conviction (3.22 times) before age 26 if the parent had a history of 4 or more ELS, compared to parents with a history of 0–1 ELS (Barnert et al., 2023). ELS experienced by a parent or grandparent may include toxins in the environment, violence in the neighborhood, and discrimination and racism (Amaro et al., 2021). These larger structural issues persist across decades while also limiting the upward mobility of the next generation, an effect known as minorities’ diminished returns (Assari, 2020).
Not all intergenerational effects are caused by external community or environmental factors. As mentioned in Section 2.6.2, genetic and epigenetic mechanisms transmit risk of depression, anxiety, and substance use across generations (Thumfart et al., 2022). In a nation-wide sample of 9–10 year old children, having both parents and grandparents with a history of depression resulted in a higher prevalence of psychiatric disorders than just having one parent with depression (Van Dijk et al., 2021). Thus, grandparent and parental experiences may encode changes in genetics, behavior, or environment that may also contribute to symptoms of depression, anxiety, and substance use.
2.6.4. Protective Factors
Protective factors and resources further modify the relationships connecting ELS and substance use. Resilience is the ability to overcome risk exposure, avoid negative trajectories associated with risk factors, and successfully cope with traumatic experiences (Fergus & Zimmerman, 2005). Factors influencing resilience can be measured using multiple scales, and increased resilience is associated with decreased cortisol levels in the face of perceived stress (Lehrer et al., 2020), lower depressive symptoms, and weaker correlations between depression and SUD (He et al., 2022). Some factors, such as self-esteem, can contribute to risk or resilience depending on whether self-esteem is low or high. Similarly, academic achievement, parental supervision, and family SES all contribute to risk when low and resilience when high. But not all factors have this scalar relationship. Involvement in extracurricular activities can be beneficial, but a lack of these activities is not necessarily representative of risk (Fergus & Zimmerman, 2005). Parental divorce, typically viewed as a risk factor, can be beneficial if it reduces daily family conflicts.
Not all experiences of stress are harmful, and adolescent adaptability can result in positive changes. Stress can even serve as an “inoculation,” presenting the opportunity to develop and use positive coping mechanisms (Figure 2) and problem-solving skills (Schiele & Domschke, 2018). Mild to moderate stress during adolescence has shown protective effects against anxiety-like behaviors in both primate and rat models (Parker et al., 2004; Rana et al., 2016). Intermittent moderate stress during adolescence was protective in primates placed into a novel environment, as previously stressed adolescent primates displayed less anxiety-like behavior and lower cortisol levels than control primates (Parker et al., 2004). Adolescent male rat models exposed to chronic mild stress scored lower than controls on measures of anxiety- and depressive-like behaviors and spent more time socializing than their non-stressed peers (Rana et al., 2016). These lower-level exposures to stress have also proven to be beneficial among human adolescents. In a longitudinal study of adolescents, Oldehinkel et al. (2014) examined the effect of ELS across the first 11 years of life on later depression onset after recent stressful life events. Adolescents who had not yet had a major depressive episode by age 16 were split into low, average, and high ELS groups and assessed for stressful life events occurring between the ages 16–19. High ELS showed a protective effect, lowering the probability of depression onset among subjects who reported a high number of stressful life events between age 16–19. Conversely, the low ELS group had the highest probability of depression onset when reporting a high number of stressful life events. From this study, we can theorize that learning and practicing appropriate coping mechanisms can help adolescents prepare for future stressors.
Fig. 2
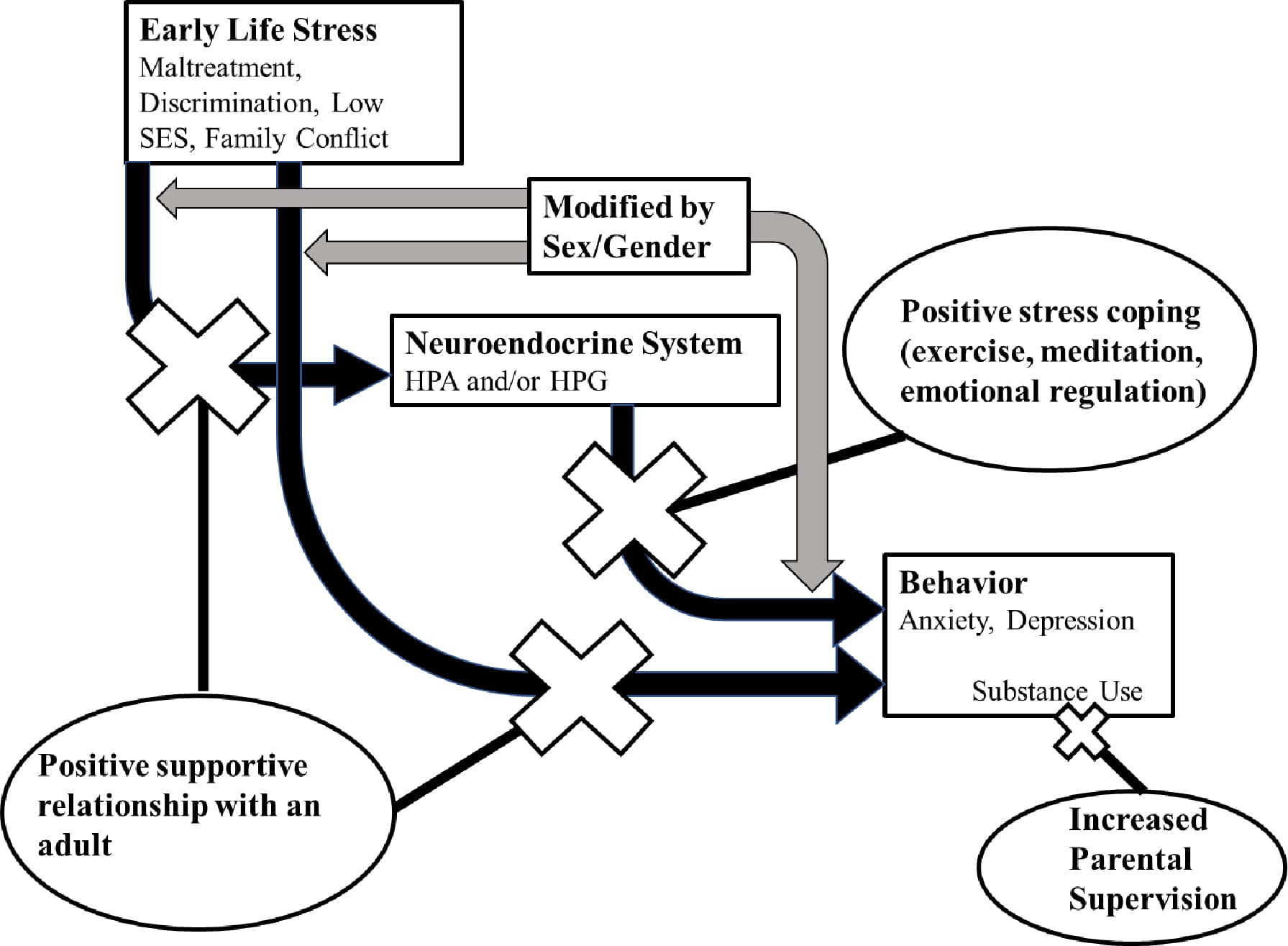
The same neuroendocrine plasticity that results in negative health effects of ELS can also assist in the recalibration of the system in response to a healthier environment. According to social support theory, friendships and positive relationships (Figure 2) can buffer the effects of ELS, decreasing risk for developing mood disorders (Cohen & Wills, 1985; Schiele & Domschke, 2018). Placement of adopted children into a supportive and well-resourced environment reverses the blunting of the HPA axis and increases normative association of cortisol and DHEA (DePasquale et al., 2019; Howland et al., 2020). Parental and family support, connectedness, and monitoring (Figure 2) are particularly powerful contributors to adolescent resilience (Aschengrau et al., 2023; Fergus & Zimmerman, 2005; Pugh & Richter, 2023). National longitudinal studies, such as the ABCD study, help identify these risk or resilience factors across various communities (Assari, 2020; Assari et al., 2020; Martz et al., 2022; Vijayakumar et al., 2023; Volkow et al., 2018; Wade et al., 2022). Diverse samples allow insight into ways in which race and ethnicity can interact with resilience factors, generating differential outcomes even given the same number of ELS (Assari, 2020). Thus, studies of resilience can help direct interventions, indicating at what level (independent, economic, community, structural) assistance can be most impactful.
3.1. Limitations to our model
Our focus was on sex and gender differences in the relationship of ELS and the endocrine system leading to internalizing problems and substance use, as depicted in Figure 1. Despite numerous studies, these domains have not been well-integrated. The focus present in our model necessitated exclusion of the myriad other factors which may influence outcomes within each domain. Specifically, papers focusing on prenatal ELS or studies of justice-system involved youth were not included within this review. Some bias in reporting may have been introduced due to the focus on internalizing disorders (depression and anxiety) rather than externalizing disorders associated with ELS. Although externalizing behaviors in both boys and girls are indicative of higher impulsivity, sensation-seeking, and enhanced response to reward (Hardee et al., 2018; Heitzeg et al., 2018), there is still a higher prevalence among girls for internalizing disorders in the presence of ELS (Chaplin et al., 2018). Yet, as adolescents transition to adulthood societal expectations surrounding gender roles can dictate how boys vs. girls are “allowed” to express emotion, seek support, and ask for help (Rice et al., 2018). These social influences can mediate choices in coping strategies, cognitive habits such as rumination, and concerns regarding peer and parental approval, which may result in differential expression of negative emotions by gender during adolescence (i.e., internalizing vs externalizing disorders) (Oldehinkel & Bouma, 2011).
4.1. Implications for Interventions and Future Research
At the school programming level, some districts are already implementing ELS interventions for young children, such as the Cognitive Behavioral Intervention for Trauma in Schools (Stein et al., 2003) or the Support for Students Exposed to Trauma (Jaycox et al., 2009). Administration of these programs by schools to youths who have experienced trauma decrease the barrier to treatment that exists for many adolescents (Jaycox et al., 2009). Additionally, the screening process helps to identify those at risk for mental health disorders before the symptoms become visible to parents and educators (Stein et al., 2003). Much work has been done on developing interventions through community and school programs to reduce long-term effects of trauma, including both interpersonal and environmental trauma, such as exposure to violence (Jaycox et al., 2014; Konanur et al., 2015). Immediately after a traumatic event, schools and communities are often willing to provide support for students, yet those who are impacted on a long-term basis (6 months to 2 years after the event) are in need of longer-term interventions (Jaycox et al., 2014).
Research-supported use of biomarkers, such as HPA and HPG axis measures, may provide an accessible, low-burden metric of proof of the need for intervention at these later dates. ELS-induced HPA and HPG axis alterations persist through adolescence and into adulthood, and providing this measure of long-term dysregulation may assist researchers in persuading funders and lawmakers to secure limited funding and resources for additional research and intervention programs. However, the use of biomarkers should be restricted to the provision of evidence of need for resources and services, rather than as a means of profiling or labeling young people. Stress management, conflict resolution, and positive health habits championed at the community and family levels may fill in the gaps in resilience observed in some adolescent populations, while also normalizing these behaviors among peers.
Future research into the pathway from ELS to depression, anxiety symptoms, and substance use should include analysis of sex/gender and HPA and HPG axis function during adolescence. The observed increase in prevalence of SUD among adult women with a comorbid mood disorder and a history of moderate-to-severe ELS (Evans et al., 2017) suggests gender differences and a common background in the etiology of these disorders. Studying relationships among ELS, mood, and substance use within a longitudinal study beginning just prior to puberty would allow researchers to track the emergence of internalizing symptoms and substance use behaviors, similar to what is seen in (Barnhart et al., 2022). Including longitudinal tracking of hormones, such as DHEA, testosterone, estradiol, and cortisol, could aid in determining the directionality of the relationships among ELS, HPA and HPG axis function, mood, and substance use. Further research may utilize imaging and behavioral data to track the neurodevelopmental impact of HPA and HPG alterations on adolescent substance use, sex and gender differences in these alterations, and their associations with ELS.
4.2. Conclusion
The path from ELS to adolescent substance use is a complex one, but sex-dependent biological pathways have been demonstrated in the research literature. ELS alters both HPA and HPG axes and these effects are stronger in females than males during puberty. This increases the risk of mood disorder symptoms and subsequent substance use in adolescent girls relative to boys. A better understanding of the sex and gender differences in the relationship between ELS and stress hormones will allow for advances in therapeutics and interventions better tailored to the differing needs of girls and boys struggling with depression, anxiety, and substance use as well as interventions to prevent their onset. Finally, outlining the biological repercussions (such as persistent alteration of HPA and HPG axis function) and associated social costs (i.e., anxiety, depression, and substance use) of structural and societal factors that contribute to ELS builds evidence to support the large-scale changes necessary to halt the continued propagation of ELS across generations.