Abstract
Adolescents' perceptions of parent and peer norms about externalizing behaviors influence the extent to which they adopt similar attitudes, yet little is known about how the trajectories of perceived parent and peer norms are related to trajectories of personal attitudes across adolescence. Neural development of midline regions implicated in self–other processing may underlie developmental changes in parent and peer influence. Here, we examined whether neural processing of perceived parent and peer norms in midline regions during self-evaluations would be associated with trajectories of personal attitudes about externalizing behaviors. Trajectories of adolescents' perceived parent and peer norms were examined longitudinally with functional neuroimaging (n = 165; ages 11–16 years across three waves; 86 girls, 79 boys; 29.7% White, 21.8% Black, 35.8% Latinx, 12.7% other/multiracial). Behavioral results showed perceived parent norms were less permissive than adolescents' own attitudes about externalizing behaviors, whereas perceived peer norms were more permissive than adolescents' own attitudes, effects that increased from early to middle adolescence. Although younger adolescents reported less permissive attitudes when they spontaneously tracked perceived parent norms in the ventromedial and medial pFCs during self-evaluations, this effect weakened as they aged. No brain–behavior effects were found when tracking perceived peer norms. These findings elucidate how perceived parent and peer norms change in parallel with personal attitudes about externalizing behaviors from early to middle adolescence and underscore the importance of spontaneous neural tracking of perceived parent norms during self-evaluations for buffering permissive personal attitudes, particularly in early adolescence.
INTRODUCTION
Adolescence is a formative period when establishing healthy attitudes lays the foundation for long-term health behaviors. Permissive attitudes about everyday externalizing behaviors (e.g., condoning health risk or antisocial behaviors) can affect an adolescent's likelihood of engaging in such behaviors (e.g., Neppl, Dhalewadikar, & Lohman, 2016). In adolescence, the perceived attitudes of parents and peers (i.e., perceived norms) are important in influencing the development of adolescents' own attitudes. Information about others' choices and outcomes, especially those of peers, has been found to selectively alter computations of risk tolerance from adolescence into early adulthood (Braams, Davidow, & Somerville, 2019, 2021; Powers et al., 2018). Compared to early adolescents and young adults, late adolescents are especially likely to make a safe choice when their peers chose the safe option but also are least likely to adjust their risk attitudes when peers make risky choices (Braams et al., 2019). Moreover, value-based computations (e.g., risk aversion) also drive preferences for prioritizing information from parents over peers (or vice versa) during decision-making. Previous studies in young adults have revealed that a preference for parent over peer influence selectively emerges in financial decisions, risk taking, and probabilistic learning, but not in representations of self–other similarity (Guassi Moreira, Tashjian, Galván, & Silvers, 2020, 2021). Conversely, decision-making becomes more equivocal (i.e., no preference between parent and peer influence) when social consequences are involved or when individuals' own attitudes conflict with both parents and peers (Guassi Moreira et al., 2021; Do, McCormick, & Telzer, 2020). Taken together, these findings underscore the age- and context-dependent nature of how parents and peers influence adolescent decision-making.
In contexts without overt pressures, adolescents tend to misestimate how permissive the attitudes are of others (Perkins, Perkins, Jurinsky, & Craig, 2019; Stattin & Kim, 2018), with such misperceptions influencing adolescents' own attitudes (DeWall, 2010). Moreover, the magnitude of perceived discrepancies between personal attitudes and those of others can directly influence the reevaluation of personal attitudes to align more closely with the perceived social norm, regardless of its accuracy (Perkins, 2002). For instance, if adolescents perceive that their parents endorse less permissive attitudes about externalizing behaviors than they do, then spontaneously (i.e., without instruction) representing the perceived norms of parents (e.g., my parent would disapprove of this externalizing behavior) during self-evaluations may reduce adolescents' likelihood of endorsing more permissive attitudes. In contrast, if adolescents perceive that their peers endorse more permissive attitudes, then representing the perceived norms of peers (e.g., my peers approve of this externalizing behavior) during self-evaluations may increase adolescents' likelihood of endorsing permissive attitudes. Given that adolescents' attitudes strongly predict their future behavior (Napper, Kenney, & LaBrie, 2015), their perceptions of parent and peer norms can have significant long-term health consequences. In the current study, we used longitudinal experimental and neuroimaging measures to characterize the neurocognitive correlates of perceived parent and peer norms that support the development of adolescents' own attitudes about externalizing behaviors.
Neural Processes Associated With The Development of Adolescent Attitudes
Developmental changes in social influence may be shaped by the spontaneous activation of the perceived norms of others during the formation of personal attitudes. This social cognitive process is likely aimed at facilitating appropriate choices when independent decision-making (i.e., based solely on self-evaluations) is still developing (Moses-Payne, Habicht, Bowler, Steinbeis, & Hauser, 2021). Indeed, children and adults have been found to spontaneously represent the mental states of others using the same brain circuitry involved in their own choices (Lim et al., 2016; Suzuki et al., 2012). Of particular interest to understanding adolescent social influence are midline cortical brain regions that undergo significant maturation during the transition to adolescence and support representations of the self and others during decision-making (Blakemore, 2012). These include brain regions implicated in representing one's own and others' thoughts (e.g., medial pFC [mPFC]; Denny, Kober, Wager, & Ochsner, 2012; Wagner, Haxby, & Heatherton, 2012), computing the value of one's own or others' attitudes (e.g., ventromedial pFC [vmPFC]; Klucharev, Hytönen, Rijpkema, Smidts, & Fernández, 2009), and monitoring inconsistencies between one's own and others' beliefs (e.g., dorsomedial pFC [dmPFC], dorsal ACC [dACC]; Izuma & Adolphs, 2013; Amodio & Frith, 2006). We hypothesize that neural sensitivity within midline cortical regions is involved in representing perceived parent and peer norms and is associated with personal attitudes about externalizing behaviors over the course of adolescence.
Self–Other Representation
The development of attitudes about externalizing behaviors is thought to involve the ability to represent one's own attitudes and the attitudes of others. The mPFC is consistently associated with understanding and representing social knowledge about one's self and of others (Wagner et al., 2012), especially mental states (e.g., thoughts; Denny et al., 2012). For example, in adolescents, the mPFC is more activated when being watched by another peer than alone (Somerville et al., 2013) and when forming self-evaluations that spontaneously integrate what others think (Van der Cruijsen, Peters, Zoetendaal, Pfeifer, & Crone, 2019). This mPFC response is elevated in adolescents relative to children or adults (Van der Cruijsen et al., 2019; Somerville et al., 2013) and is positively associated with changes in risk attitudes in response to peer influence (Braams et al., 2021). These data suggest that mPFC activation may be especially crucial to adolescents' representation of others' attitudes when constructing their own attitudes about externalizing behaviors.
Valuation
Adolescents may redefine their own attitudes as they learn about the perceived attitudes of their parents and peers, a process that involves computing and comparing the value of their own attitudes and the attitudes of others. The vmPFC, a key region for value computations, tracks the relative value of individuals' and others' attitudes that are similar to prediction errors in reinforcement learning (Klucharev et al., 2009), which signal the need to adjust behavior in the future. Adolescents who exhibit greater vmPFC activation when viewing peer ratings are more likely to align their attitudes with those of their peers (Welborn et al., 2016; Cascio, O'Donnell, Bayer, Tinney, & Falk, 2015), a relationship further moderated by the specific behavior under consideration (Do et al., 2020). Furthermore, a recent study found the extent to which the brain represents the relative value between two close others (one's parent vs. friend) predicts stronger preferences for the higher value source during decision-making (Guassi Moreira et al., 2023). Taken together, these findings highlight the role of value-based processing in adolescents' motivation to align their attitudes toward the attitudes of others.
Social Monitoring
To determine whether it is better to make decisions independently or in consideration of others' attitudes about externalizing behaviors, people must monitor and respond to discrepancies in self- and other-related representations (Suzuki, Adachi, Dunne, Bossaerts, & O'Doherty, 2015). The dmPFC is involved in representing the mental states of others (Amodio & Frith, 2006) and, together with the closely adjacent dACC, tracks cognitive inconsistencies between one's own and others' preferences (Izuma & Adolphs, 2013). For example, dmPFC and dACC activity increase when individuals' opinions conflict versus match with the group opinion, which predicts subsequent behavioral adjustments (Klucharev et al., 2009). These findings suggest that the development of attitudes about externalizing behaviors may be guided by the extent to which one's own preferences, and the degree to which it deviates from those of others, are represented in the dmPFC and dACC.
Current Study
Most previous research has examined neural mechanisms of social influence in the presence of external pressures (Telzer, Rogers, & van Hoorn, 2017) and age differences in social influence between individuals using cross-sectional designs (Knoll, Leung, Foulkes, & Blakemore, 2017). Few studies have used longitudinal designs to examine within-person changes (i.e., year-to-year changes for a given adolescent) or examined spontaneous tracking of perceived social norms to implicitly assess how an adolescent incorporates others' perspectives in decision-making. Using longitudinal neuroimaging in a diverse sample spanning early to middle adolescence, the current study tested two preregistered hypotheses regarding how adolescents' own attitudes about externalizing behaviors develop in relation to their perceived parent and peer norms at the behavioral and neural levels: osf.io/mbucn/. During annual fMRI scans across 3 years, participants rated how good or bad they thought externalizing behaviors were (Self condition), as well as the perceived norms of their primary guardian (Parent condition) and peers their age (Peers condition). Our preregistered hypotheses included two aims:
Our first aim was to examine how adolescents' own attitudes about externalizing behaviors develop in parallel with their perceived parent and peer norms from early to middle adolescence. Previous work has shown that youth often misestimate the permissiveness of others' attitudes (Borsari & Carey, 2003). Building off this literature, we predicted that self–other discrepancies in attitudes would change over time, aligning with the presumed shift from the importance of parental-to-peer influence during adolescence (Nelson, Jarcho, & Guyer, 2016). Behaviorally, we expected that trajectories of adolescents' own attitudes would become increasingly permissive, such that they would rate externalizing behaviors as less bad over time. Moreover, we hypothesized that such trajectories of adolescents' own attitudes would diverge more from perceived parent norms over time (i.e., increasing self-parent discrepancies with age) and diverge less from perceived peer norms over time (i.e., decreasing self-peer discrepancies with age).
When forming their own attitudes, adolescents not only represent their self-evaluations but may also spontaneously activate the perceived norms of their parents or peers for the same behavior (Lim et al., 2016; Suzuki et al., 2012). Our second aim was to examine how longitudinal trajectories of adolescent attitudes are influenced by spontaneous neural representations of perceived parent and peer norms in midline cortical brain regions (mPFC, vmPFC, dmPFC, dACC) during self-evaluations of externalizing behaviors. At the neural level, we hypothesized that spontaneous (i.e., without instruction) tracking of perceived parent and peer norms during self-evaluations in midline cortical regions would be associated with trajectories of adolescents' own attitudes about externalizing behaviors. Crucially, we expected this brain–behavior association would change as adolescents grew older and spontaneously activated the perceived norms of different reference groups during decision-making (parents vs. peers). Insofar as the impact of the perceived norms of parents versus peers shifts in importance from early to middle adolescence, we predicted that adolescents would either consistently or decreasingly track their perceived parent norms when evaluating their own attitudes as they aged. This effect would imply that adolescents either consistently or decreasingly rely on perceived parent norms to guide their personal attitudes about externalizing behaviors across adolescence. In contrast, we predicted that adolescents would increasingly track their perceived peer norms when evaluating their own attitudes as they aged, suggesting an increasing reliance on perceived peer norms to guide personal attitudes from early to middle adolescence.
METHODS
Participants
Two cohorts, each separated by a 1-year interval, were recruited from a larger study of 873 sixth and seventh grade students in the Southeast United States to meet a target sample size of 150 participants, which was determined by a power analysis performed during grant submission (before data collection) that accounts for expected attrition across the 3-year study. Adolescent participants were initially screened by phone using the following exclusion criteria: not fluent in English, history of neurological or learning problems, and presence of MRI contraindications (e.g., metal in body, claustrophobia). Participants who currently used psychoactive medications for attention-deficit/hyperactivity disorder were instructed to abstain for 24 hr before the visit. The first cohort of 148 participants was recruited at Wave 1, but eight participants met exclusion criteria at their baseline visit and were subsequently excluded and not recruited at the follow-up waves: One was not fluent in English, five were unable to complete the scan (e.g., claustrophobia, noncompliant), one had a metal implant, and one had a brain abnormality. Of the eight participants excluded at baseline, three participants completed the task behaviorally and were retained for behavioral analyses, leaving a final sample of 143 participants at the first wave (Mage = 12.8 years, SD = 0.52 years, range = 11.93–14.49 years; 73 girls, 70 boys, 0 other). Given the expected attrition and excluded participants at Wave 1, a second cohort comprising 30 participants was recruited at Wave 2 to meet recruitment goals (Mage = 13.30 years, SD = 0.64 years, range = 12.42–14.52 years; 18 girls, 12 boys, 0 other), drawn from the same study as the first cohort. Note that the second cohort could only contribute a maximum of two waves of data within the 3-year study period.
In total, data were collected from 173 participants across both cohorts. Eight participants were excluded after data quality checks (e.g., no variability in ratings across trials/conditions), leaving a final sample of 165 participants included in analyses. Across the 3-year study period, participants completed up to three annual fMRI scans, which included a baseline scan starting when they were 11–14 years old and follow-up assessments approximately every 12 months thereafter. Participants who were unable to do follow-up scans because of new exclusion criteria (e.g., braces) completed the task behaviorally outside the scan. In total, we had 385 behavioral and 371 fMRI data points. At baseline (i.e., first year of participation), the age and sex distribution were M = 12.94 years, SD = 0.57 years, and range = 11.93–14.52 years, with 86 girls (52.1%), 79 boys (47.9%), 0 other sex (0%). The race/ethnicity distribution was as follows: White (n = 49, 29.7%), Black (n = 36, 21.8%), Latinx (n = 59, 35.8%), and other/multiracial (n = 21, 12.7%). The median educational level of the primary guardian/parent was some college (n = 51, 30.9%), with a range of less than eighth grade (n = 17, 10.3%) to a professional degree (e.g., MD; n = 3, 1.8%). Adolescents and their primary caregiver provided written assent and consent and were compensated for their time.
Attitude Evaluation Task
During fMRI, participants completed the attitude evaluation task, which assesses adolescents' own attitudes about externalizing and perceptions of their parents' and their peers' norms regarding the same behaviors. Behaviors were selected based on a prior study examining parent and peer influence on attitude change using a similar age range (n = 39; 12–14 years; (Do et al., 2020). From this independent sample, we selected 48 externalizing behaviors (e.g., risk taking, antisocial behavior; Table 1) based on well-validated and common measures of youth behaviors (e.g., Child Behavior Checklist) and with relatively high response variability to reflect real-life individual differences in adolescent attitudes. In the attitude evaluation fMRI task, participants were presented with 48 externalizing behaviors and instructed to use button boxes to rate how bad or good they thought each behavior was on a 10-point Likert scale (0 = very bad, 9 = very good; Figure 1). The behaviors were presented in a random order for 5 sec and separated by a jittered intertrial interval (range = 1.3–3.4 sec, M = 2.2 sec). A “Too Late!” message (1 sec) was displayed if the participant did not rate the behavior within the allotted time. The task was self-paced, such that the screen advanced to the next trial upon participant response or the “Too Late!” message.

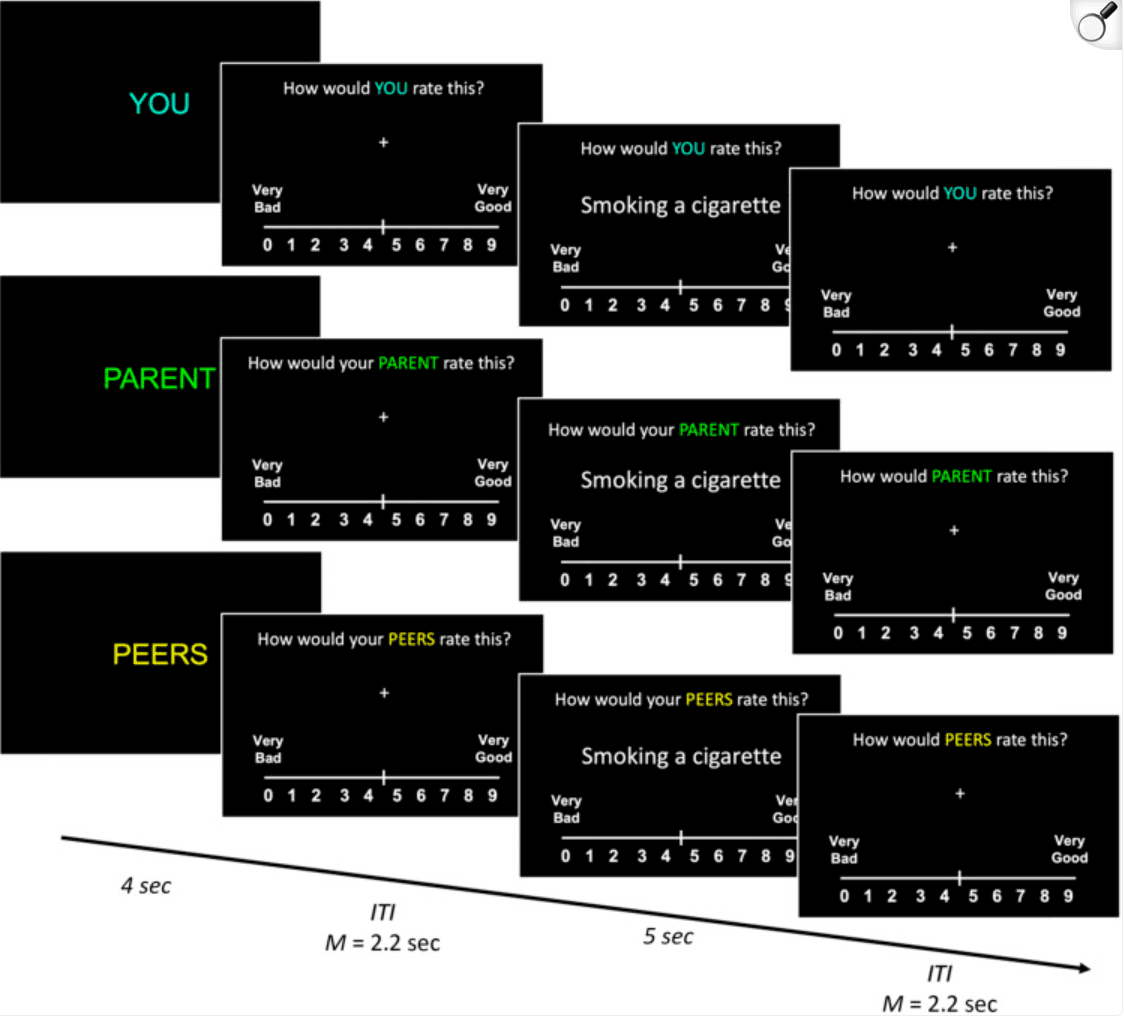
Figure 1: Schematic illustration of attitude evaluation task. ITI = intertrial interval. The attitude evaluation task had three conditions: Self, Parent, and Peers (Figure 1). In the Self condition, participants rated how good or bad they thought each behavior was. To assess adolescents' perceptions of parent and peer norms, participants also rated how good or bad they thought their primary guardian (Parent condition) and other teenagers around the same age (Peers condition) would perceive the same behavior. Each condition was composed of four blocks of 12 behaviors (trials) each, resulting in 48 trials per condition. Before each condition, participants were presented with an initial instruction cue for 4 sec indicating the condition type (Self, Parent, Peers) of the subsequent block of trials. Participants completed two runs lasting up to 8 min each. Each run was composed of six blocks, with the conditions for each block presented in a random order. In total, the task included 144 trials across both runs, with 72 trials per run.
fMRI Data Acquisition, Processing, and Analysis
Imaging data were collected using a 3-T Siemens Prisma MRI scanner with a 32-channel head coil. For registration purposes, anatomical scans were acquired using a T1* magnetization-prepared rapid-acquisition gradient echo (repetition time [TR] = 2.4 sec, echo time [TE] = 2.2 msec, flip angle = 8°, field of view [FOV] = 256 mm, slice thickness = 0.8 mm, 208 slices, voxel size = 0.8 × 0.8 × 0.8) and a T2*-weighted, matched-bandwidth, high-resolution, anatomical scan (TR = 5.7 sec, TE = 65 msec, flip angle = 120°, FOV = 230 mm, slice thickness = 3 mm, 38 slices, voxel size = 1.2 × 1.2 × 3 mm3). BOLD EPI were acquired using a T2*-weighted gradient echo sequence (TR = 2 sec, TE = 25 msec, flip angle = 90°, FOV = 230 mm, slice thickness = 3 mm, 37 slices, voxel size = 2.5 × 2.5 × 3 mm3). Matched-bandwidth and EPI scans were obtained using an oblique axial orientation to maximize brain coverage and reduce signal dropout.
Imaging data were preprocessed using FMRI Expert Analysis Tool Version 6, part of the FMRIB Software Library (Smith et al., 2004). The following prestatistics preprocessing was applied: motion correction using MCFLIRT (Jenkinson, Bannister, Brady, & Smith, 2002), skull-stripping using BET (Smith, 2002), and spatial resampling to 2 mm. A registration matrix was estimated between functional images, high-resolution structural images, and standard Montreal Neurological Institute 2-mm brain using FLIRT (Jenkinson et al., 2002; Jenkinson & Smith, 2001). Spatial smoothing with a 6-mm Gaussian kernel of FWHM was then applied. To further correct for motion and physiological noise in the imaging data, we conducted an independent component analysis denoising procedure using MELODIC and an automated signal classification toolbox (classifier NP threshold = 0.3; Tohka et al., 2008).
Individual-level Analysis
Individual-level, fixed effects analyses were conducted on the preprocessed imaging data using general linear model (GLM) in the SPM software package (SPM12; Wellcome Department of Cognitive Neurology). The experimental task was modeled as an event-related design. In each GLM, the regressor of interest included decisions with a duration equal to the RT of that choice, modeled separately for each condition (Self, Parent, and Peers).
All analyses in the current study focused on neural activation during the Self condition during which participants evaluated their own ratings. We chose to focus on neural activation during the Self condition as it tracks self, parent, or peer ratings to implicitly assess whether adolescents spontaneously (i.e., without instruction) represent and incorporate their parents' or peers' ratings in their own evaluations. We ran three separate GLMs on the Self condition to test how strongly brain activation is correlated with trial-level fluctuations in (1) their own self-evaluations, (2) the perceived norms of their parent on the same behavior, and (3) the perceived norms of their peers on the same behavior, all of which came from their in-scanner ratings during the attitude evaluation task. Neural activation during the Parent and Peers conditions is not included in this study because these conditions would assess neural differences during explicit (rather than spontaneous) mentalizing about perceived parent and peer norms. We first examined parametric modulation of brain activation by the strength of self-evaluations (GLM-1). For each participant, adolescents' own rating on each trial (from the Self condition) was centered relative to their minimum rating across 48 behaviors and entered as a parametric modulator at the time of their own decision (Self condition). Parameter estimates from GLM-1 represent how strongly activation in brain regions scaled with increases in self-evaluations at the time of adolescents' own decision (i.e., “neural tracking of self-evaluations”). To test whether the brain also spontaneously tracks the norms of others while adolescents evaluated their own attitudes (i.e., Self condition), we examined parametric modulation of brain activation by adolescents' perceptions of their parent norms (GLM-2) and their peer norms (GLM-3). GLM-2 and GLM-3 sought to examine whether adolescents' perceptions of parent and peer norms mapped onto the neural tracking of how they personally thought about the externalizing behavior. Because any information about the perceived norms of others while adolescents evaluated their own attitudes would be retrieved spontaneously (i.e., without instruction during the Self condition), and trial-wise behaviors were matched across conditions, these analyses examined perceived parent (or peer) norms on the same behavior in which adolescents were representing neurally. For each participant, the perceived parents' rating for a matched behavior (from the Parent condition) or the perceived peers' rating for a matched behavior (from the Peer condition) was centered relative to the minimum parents' or peers' ratings across 48 behaviors and entered as a parametric modulator at the time of adolescents' own decision on the same behavior (Self condition). Parameter estimates from GLM-2 represent how strongly brain activation correlated with perceived parent norms on the same behaviors in which adolescents were personally evaluating (i.e., “neural tracking of parent norms”), and parameter estimates from GLM-3 represented how strongly brain activation correlated with perceived peer norms in which adolescents were personally evaluating (i.e., “neural tracking of peer norms”). Across all GLMs, a positive parameter estimate reflects greater tracking of permissive attitudes (i.e., positive correlation between brain activation and perceived norms), whereas a negative parameter estimate reflects greater neural tracking of restrictive attitudes (i.e., negative correlation between brain activation and perceived norms).
For all three GLMs, instruction cues (fixed duration of 4 sec), error trials (e.g., missing responses), and high motion volumes (> 0.9-mm framewise displacement) were modeled as separate nuisance regressors. One participant whose functional runs at Wave 1 exceeded > 0.9-mm framewise displacement on more than 25% trials was excluded from fMRI analyses at W1 for excessive motion but retained at Wave 2 and Wave 3 because they met the motion cutoff. In addition, six motion regressors were modeled as covariates of noninterest to control for head movement. The jittered intertrial intervals were not explicitly modeled and thus served as an implicit baseline. All regressors were convolved with a canonical hemodynamic response function. High-pass temporal filtering with a 128-sec cutoff was applied to remove low-frequency drift in the time series. Serial autocorrelations were estimated with a restricted maximum likelihood algorithm with an autoregressive model order of 1. For each GLM, contrasts were generated at the individual level for the parametric regressor of the Self condition.
ROI Analysis
At the group level, we extracted activation from four a priori ROIs, which were identified based on their roles in self–other, valuation, and social monitoring processes during social decision-making: mPFC, vmPFC, dACC, and dmPFC. Bilateral ROIs were functionally defined based on prior studies of adolescent social sensitivity (dACC from Falk et al., 2014) and the Saxe Lab theory of mind atlas (mPFC, vmPFC, and dmPFC from Dufour et al., 2013). ROI masks are available on NeuroVault: neurovault.org/collections/GKSJTTUQ/.
Analysis Plan
Longitudinal Trajectories of Adolescents' Own Attitudes and Perceptions of Parent and Peer Norms
Multivariate growth model.
Our first aim was to examine whether adolescents' own attitudes about externalizing behaviors diverged more from their perceived parent or peer norms from early to middle adolescence. Preliminary analyses using univariate growth models were conducted to determine the best-fitting trajectory of age-related change (i.e., linear or quadratic) for (1) adolescents' own attitudes, (2) perceived parent norms, and (3) perceived peer norms. Each of these outcome variables were calculated as the mean rating of the 48 trials from the in-scanner ratings during the Self condition (i.e., self attitudes), the Parent condition (i.e., perceived parent norms), and the Peers condition (i.e., perceived peer norms), with higher scores reflecting more permissive attitudes. We then combined the three univariate growth models into a single multivariate growth model to examine how trajectories of adolescents' own attitudes covaried with trajectories of perceived parent and peer norms at the individual level. Specifically, we tested how much adolescents' own attitudes diverged from the perceived norms of their parents or their peers, both in terms of starting points (i.e., difference between intercepts) and rates of change (i.e., difference between slopes). Following statistical procedures for estimating growth processes in multiple outcomes simultaneously (Curran, McGinley, Serrano, & Burfeind, 2012), we fitted a multivariate growth model under full maximum likelihood to a single stacked outcome that contained repeated assessments on self, parent, and peer over time, with observations (i ≤ 9 [3 targets × up to 3 time points]) nested within individuals (j = 165). As shown in Equation 1, predictors of interest included target (dummy-coded variables that represent the intercept of each of the three outcomes: Self, Parent, Peer), continuous age (centered at minimum), and interactions between target and age (represent the slope or trajectory of each of the three outcomes: Age × Self, Age × Parent, Age × Peer). The best-fitting multivariate growth model included a random intercept and slope of each outcome to allow for individual differences in starting points and rates of change in adolescents' own attitudes and their perceived parent and peer norms. We specified a heteroscedastic residual variance structure for all three outcomes as well as a heterogeneous autoregressive error structure to allow for correlated residuals across outcomes within each time point.
Equation 1:
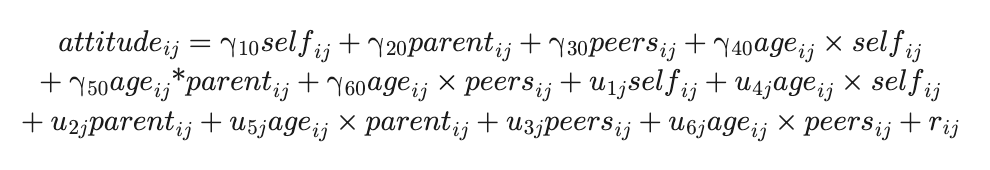
Note that multivariate growth models typically do not include an overall intercept but model the intercept (i.e., starting points:γ10selfij, γ20parentij, γ30peersij) and slope (i.e., rates of change: γ40ageij × selfij, γ50ageij × parentij, γ60ageij × peersij) of each outcome simultaneously. To control for potential sex and racial differences in trajectories of attitudes, we included dummy-coded sex (1 = female, 0 = male) and race (Black, Latinx, and other/multiracial, with White omitted as the reference group) as covariates of the intercept and slope of attitudes in the best-fitting model (not shown in [1]).
Is Neural Tracking of Parent and Peer Norms Associated with Longitudinal Changes in Adolescents' Own Attitudes?
Our second aim was to examine whether the neural correlates of perceived parent and peer norms are associated with longitudinal changes in adolescents' own attitudes about externalizing behaviors. To examine these brain–behavior associations, we tested how adolescents' own attitudes were predicted by midline cortical brain regions that track self-evaluations and perceived parent and peer norms at the time of adolescents' choice. To assess the specificity of the implicated brain regions, we fitted a separate linear mixed-effects model for each ROI to adolescents' own attitudes about externalizing behaviors, with time points (i ≤ 3) nested within individuals (j = 165). The outcome variable, adolescents' own attitudes, was calculated as the mean rating of the 48 trials from the in-scanner ratings during the Self condition. As shown in Equation 2, predictors of interest included neural tracking of each target's attitudes (self, parent, peers) as time-varying covariates, continuous age (centered at minimum), and interactions between neural tracking of each target's attitudes and age. One participant with extremely low (2 SDs below normal range) neural tracking of self-evaluations across all four ROIs at Wave 3 was winsorized for analyses. The best-fitting model included a random intercept (u0j) and slope of adolescents' own attitudes (u4jageij) to account for variability in starting points and rates of change in attitudes, respectively. We did not specify additional random effects because there would be fewer observations (≤3 time points) than random effects across participants, reducing model parsimony.
Equation 2:
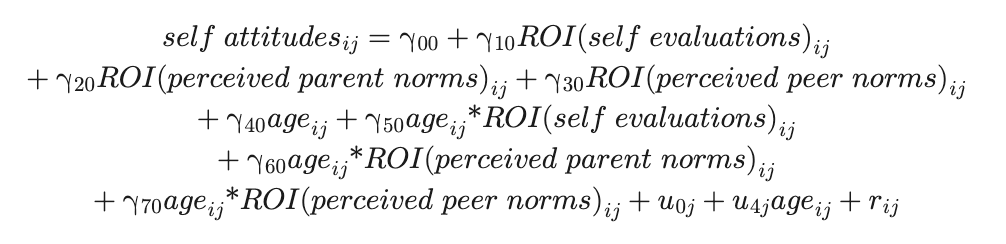
To better understand how these brain–behavior associations change from early to middle adolescence, we parsed the within-person effects (i.e., year-to-year variability) and between-person effects (i.e., between-person differences) of all ROI predictors. It is important to disaggregate within-person and between-person effects to infer whether brain–behavior associations hold within persons relative to one's own average (i.e., within-person effect) and across persons relative to the sample's average (i.e., between-person effect; Curran & Bauer, 2011). The cluster-level variable (participants' ROI) was grand mean-centered to capture person-level means (i.e., between-person effects), and the repeated measures were group-mean centered within participants to capture year-to-year deviations around person-level means (i.e., within-person effects; not shown in Equation 2). To control for potential sex and racial differences in brain–behavior associations, we included dummy-coded sex (1 = female, 0 = male) and race (Black, Latino, and other/multiracial, with White omitted as the reference group) as covariates of the intercept and slope of self-attitudes in the best-fitting model (not shown in Equation 2). We used a Benjamini–Hochberg false discovery rate correction to control for multiple comparisons across the four planned brain–behavior models (false discovery rate corrected p < .016).
RESULTS
Longitudinal Trajectories of Adolescents' Attitudes and Perceptions of Parent and Peer Norms
Descriptive statistics of attitudes about externalizing behaviors by time point and target are reported in Table 2. On average, adolescents perceived their parents to endorse less permissive norms than their own (i.e., perceived parents as less approving than they are), whereas they perceived their peers to endorse more permissive norms than their own (i.e., perceived peers as more approving than they are). Preliminary univariate growth models revealed the trajectories of each target's attitudes about externalizing behaviors were best explained by linear age functions (also see Table 3). As shown in Figure 2, adolescents on average reported increasingly permissive attitudes as they aged and perceived increasingly permissive norms from their parents and peers as they aged. The multivariate growth model revealed that adolescents' own attitudes and perceptions of parent and peer norms track together over time at the individual level. This was evidenced by significant and positive covariances among intercepts (i.e., initial levels) and slopes (i.e., rates of change) between self-attitudes and perceived parent and peer norms about externalizing behaviors (Table 4).
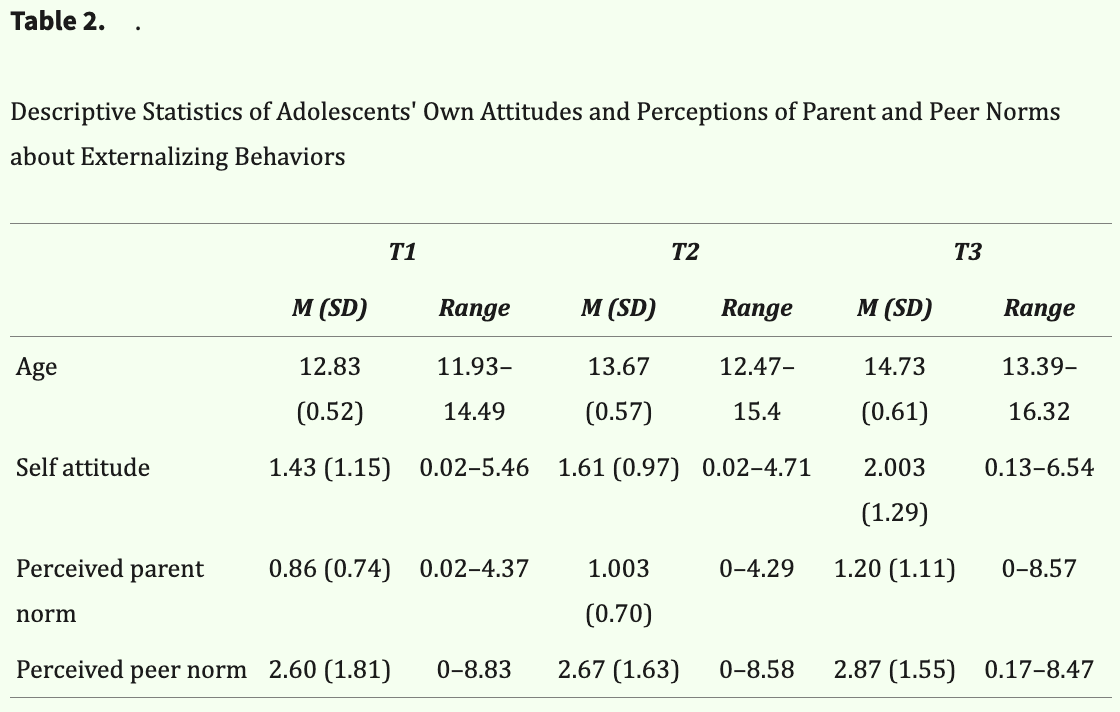
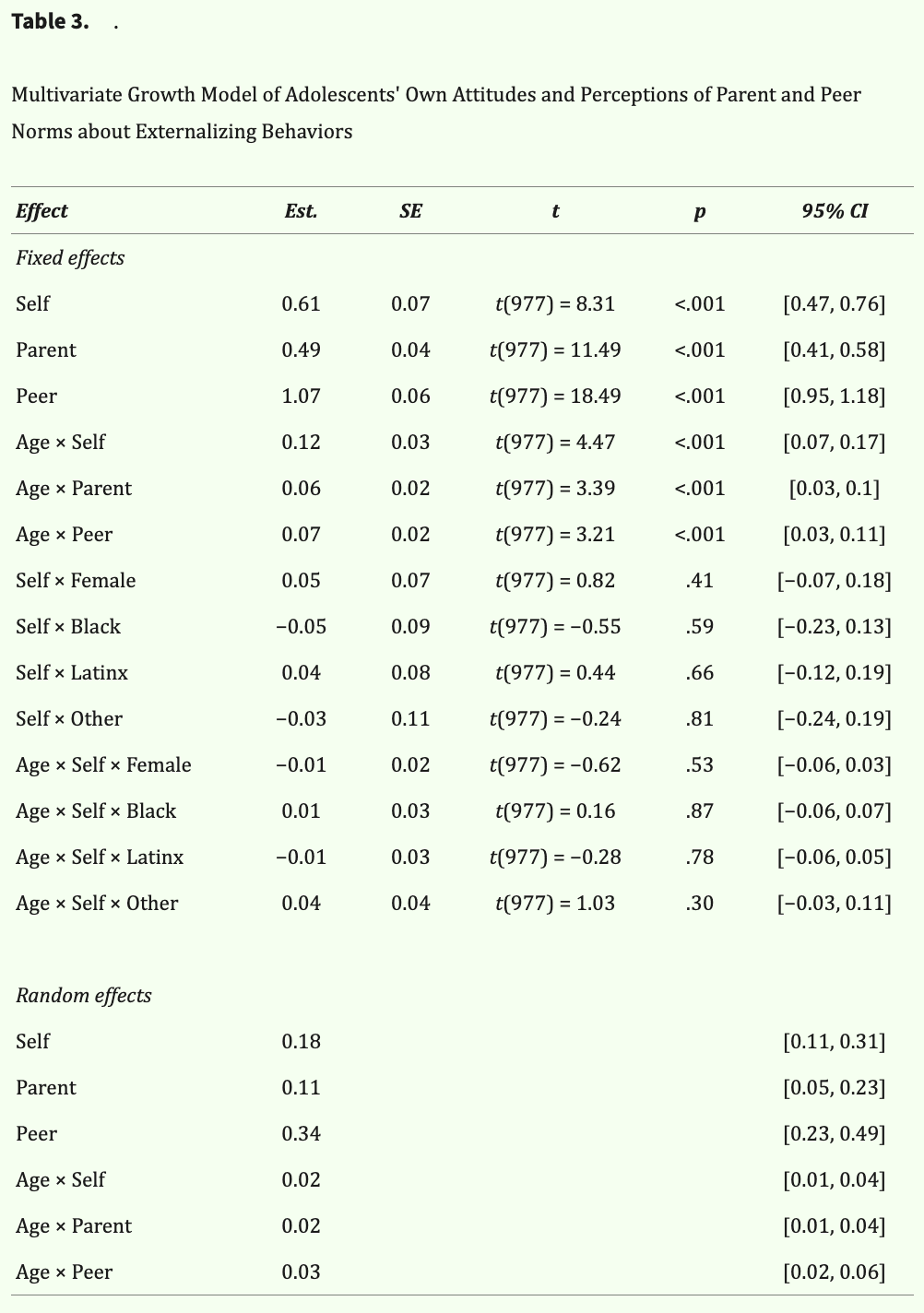
Table 3: Age was centered at the minimum age of 11 years. Sex (1 = female, 0 = male) and race (dummy-coded with “White” omitted as the reference group) were included as covariates of the intercept and slope of adolescents' own attitudes about externalizing behaviors (i.e., Self). Est = regression coefficient; SE = standard error; CI = confidence interval.
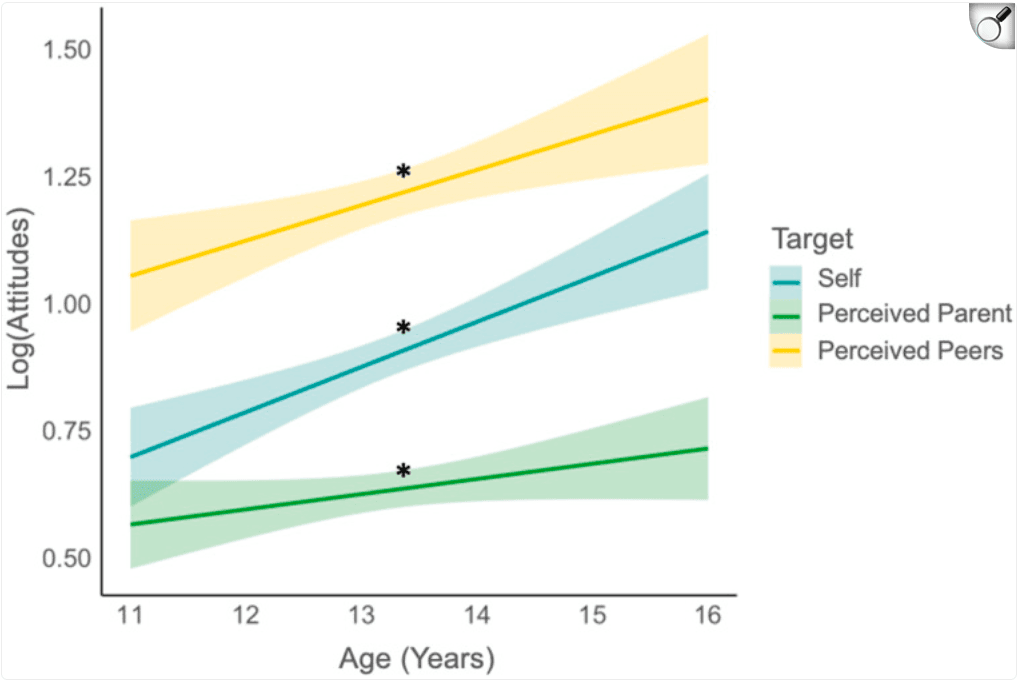
Figure 2: Trajectories of adolescents' own attitudes and the perceptions of parent and peer norms about externalizing behaviors. Adolescents' own attitudes about externalizing behaviors diverged more from their perceived parent norms over time and diverged less from their perceived peer norms over time. Simple slopes: *95% CI did not include 0.
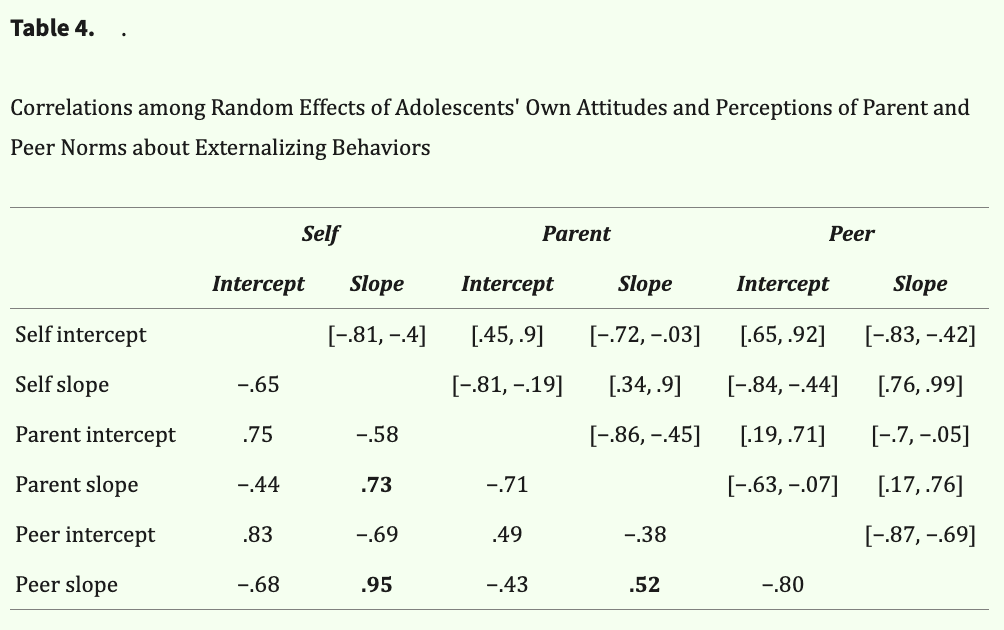
Table 4: Values below the diagonal represent correlations among random effects, whereby positive values indicate positive correlations between random effects, and negative values indicate negative correlations between random effects. Values above the diagonal represent 95% confidence intervals of correlations among random effects. Correlations among slopes of self, perceived parent, and perceived peer norms are bolded, indicating how each target's norms about externalizing behaviors co-develop over time at the individual level.
To examine whether adolescents' own attitudes about externalizing behaviors diverged from the perceived norms of their parents or peers as they grew older, we next compared the simple slopes and simple intercepts between self-attitudes and perceived parent and peer norms. Planned pairwise contrasts of simple slopes revealed significant differences between trajectories of self-attitudes and perceived parent norms [difference between slopes: F(1, 977) = 5.32, p = .02, partial η2 = .01]. To unpack these differences in trajectories, we probed the simple intercepts in early adolescence (age 12) and mid-adolescence (age 15). In early adolescence, adolescents' own attitudes significantly differed from perceived parent norms [age 12: F(1, 977) = 12.61, p < .001, partial η2 = .01], a discrepancy that significantly increased by mid-adolescence [age 15: F(1, 977) = 31.85, p < .001, partial η2 = .03]. Thus, adolescents' own attitudes and their perceived parent norms became increasingly divergent over time (Figure 2).
We also found significant differences between trajectories of adolescents' own attitudes and perceived peer norms about externalizing behaviors [difference between slopes: (F(1, 977) = 4.37, p = .04, partial η2 = .004]. Adolescents' own attitudes were significantly different from their perceived peer norms in early adolescence [age 12: F(1, 977) = 51.32, p < .001, partial η2 = .05], but this discrepancy reduced and remained significant by mid-adolescence [age 15: F(1, 977) = 16.30, p < .001, partial η2 = .02]. Thus, adolescents' own attitudes and perceived peer norms diverged less over time (Figure 2).
Is Neural Tracking of Perceived Parent and Peer Norms Associated with Longitudinal Changes in Adolescents' Own Attitudes?
Next, we investigated whether the neural correlates of perceived parent and peer norms are associated with adolescents' own attitudes about externalizing behaviors and how such brain–behavior associations change with age. Separate linear mixed-effects models for each ROI revealed neural tracking of self-evaluations and perceptions of others' norms in the mPFC and vmPFC are associated with within-person changes and between-person differences in adolescents' own attitudes (Table 5). The effects in the dACC and dmPFC trended in the same direction (ps < .08). Below, we unpack the effects for each ROI.
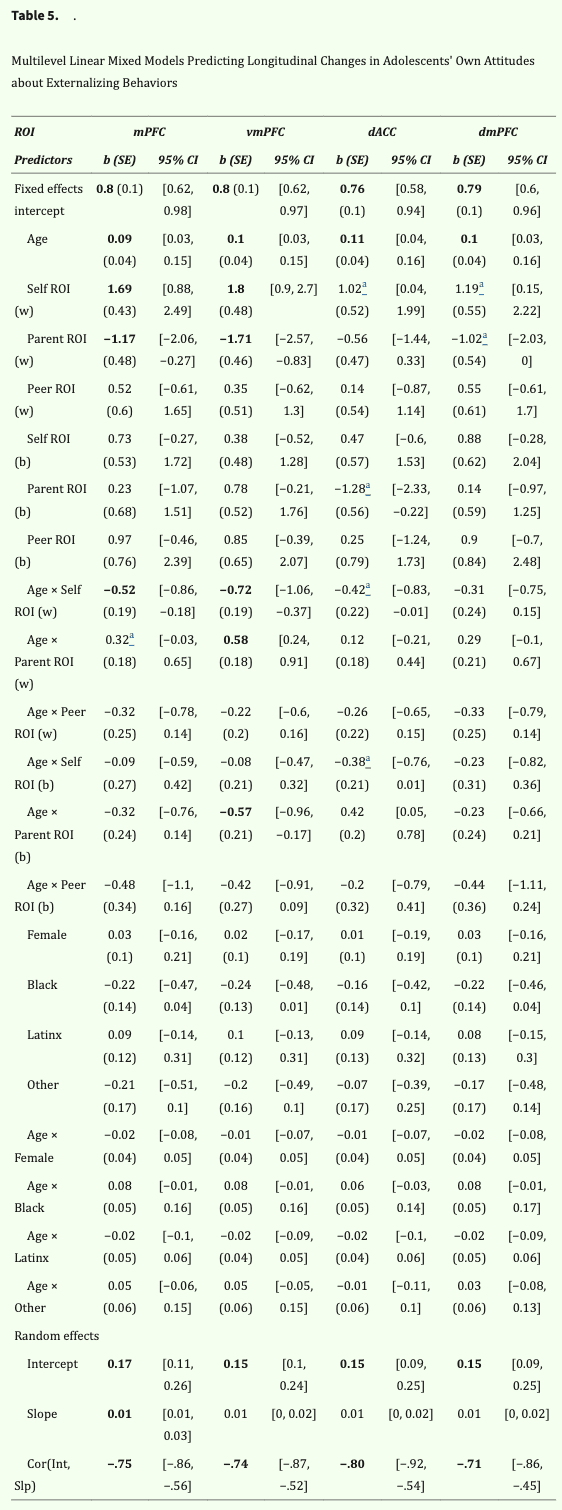
Table 5: Significant effects are bolded (i.e., 95% CI did not include 0, false discovery rate corrected). Age was centered on the minimum age of 11 years. Sex (1 = female, 0 = male) and race (dummy-coded with “White” omitted as the reference group) were included as covariates of the intercept and slope of adolescents' own attitudes about externalizing behaviors. SE = standard error; ROI (w) = within-person effect of ROI on the development of adolescents' own attitudes (i.e., across-year differences within a given person); ROI (b) = between-person effect of ROI on the development of adolescents' own attitudes (i.e., across-person differences); Cor(Int, Slp) = correlation between random intercept and random slope.
a Trending effect at more liberal thresholds (ps < .02–.08).
mPFC
Neural tracking of self-evaluations.
The linear mixed-effects model revealed a significant within-person effect of mPFC tracking of self-evaluations and its interaction with age. This indicates that, within a given individual, greater mPFC tracking of self-evaluations was associated with more permissive personal attitudes about externalizing behaviors, but this brain–behavior association weakened as the adolescent aged. To explore this interaction with age, we computed simple slopes of the within-person effect of mPFC tracking of self-evaluations in early adolescence (age 12) and mid-adolescence (age 15). As shown in Figure 3A, a younger adolescent (age 12) who exhibited greater mPFC tracking of self-evaluations was more likely to report permissive attitudes overall (age 12: b = 1.17, SE = 0.27, 95% CI [0.62, 1.71]). However, this effect went away as the adolescent aged (age 15: b = −0.39, SE = 0.40, 95% CI [−1.18, 0.39]). There was no between-person effect of mPFC tracking or its interaction with age, suggesting that age differences in adolescents' own attitudes were not moderated by mPFC sensitivity.
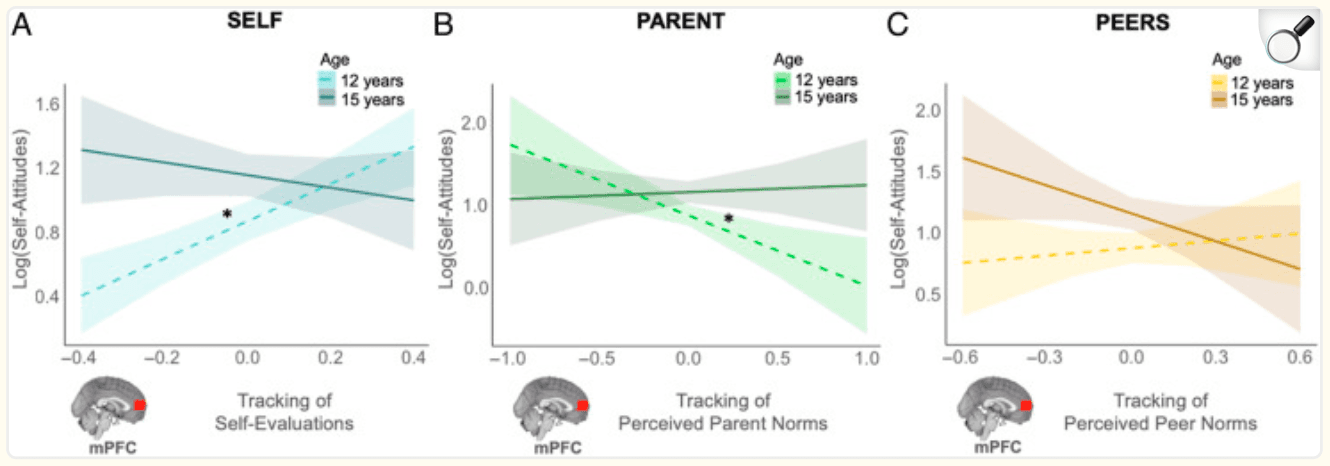
Figure 3: mPFC tracking of each target's norms is associated with longitudinal changes in adolescents' own attitudes about externalizing behaviors. Within a given adolescent, the association between mPFC tracking of each target's norms on adolescents' own attitudes about externalizing behaviors changes as they age (within-person effect). (A) A younger adolescent (age 12) who exhibited greater mPFC tracking of self-evaluations tended to endorse more permissive attitudes; this effect went away as they grew older (age 15). (B) A younger adolescent (age 12) tended to report less permissive attitudes when they exhibited greater mPFC tracking of perceived parent norms at the time of adolescents' choice; this effect went away as they grew older (age 15). (C) mPFC tracking of perceived peer norms at the time of adolescents' choice was not associated with adolescents' own attitudes. Simple slopes: *95% CI did not include 0.
Neural tracking of perceived parent and peer norms.
We found a significant main effect of mPFC tracking of perceived parent norms at the within-person level, supporting our hypothesis that an adolescent spontaneously represents their perceived parent norms when forming their own attitudes about the same behaviors. Specifically, an adolescent who exhibited greater mPFC tracking of perceived parent norms tended to report less permissive attitudes about externalizing behaviors. A trending interaction with age (p = .08) suggests that this brain–behavior association weakened as the adolescent grew older. As shown in Figure 3B, simple slopes analyses revealed that a younger adolescent (age 12) who exhibited greater mPFC tracking of parent norms was less likely to report more permissive attitudes (age 12: b = −0.85, SE = 0.31, 95% CI [−1.46, −0.25]); this effect went away as the adolescent progressed into mid-adolescence (age 15: b = 0.08, SE = 0.29, 95% CI [−0.49, 0.66]). There was no between-person effect of mPFC tracking or its interaction with age. Contrary to hypotheses, we did not find a main effect of mPFC tracking of perceived peer norms or its interaction with age at the within-person (Figure 3C) or between-person levels.
vmPFC
Neural tracking of self-evaluations.
The linear mixed-effects model revealed a significant within-person effect of vmPFC tracking of self-evaluations and its interaction with age (Table 5). To unpack the interaction, we probed the simple slopes of vmPFC tracking of self-evaluations in early adolescence (age 12) and mid-adolescence (age 15). As shown in Figure 4A, a younger adolescent who exhibited greater vmPFC tracking of self-evaluations was more likely to report permissive attitudes (age 12: b = 1.09, SE = 0.32, 95% CI [0.46, 1.71]). However, the direction of this brain–behavior association flipped as the adolescent aged, such that an older adolescent who exhibited greater vmPFC sensitivity to self-evaluations tended to report less permissive attitudes (age 15: b = −1.05, SE = 0.36, 95% CI [−1.75, −0.35]). There were no significant between-person effects.
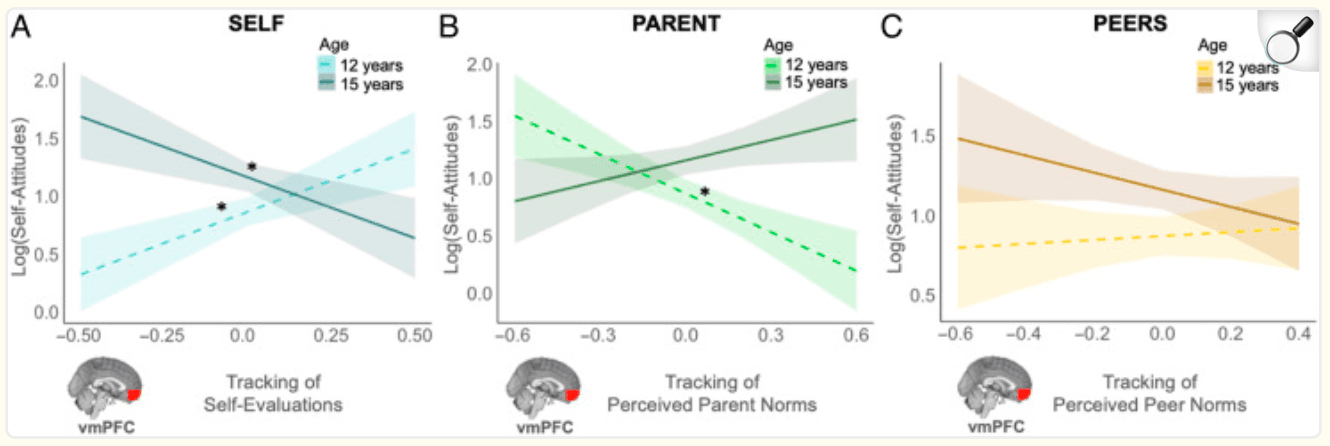
Figure 4: vmPFC tracking of each target's norms is associated with longitudinal changes in adolescents' own attitudes about externalizing behaviors. Within a given adolescent, the association between vmPFC tracking of each target's norms on adolescents' own attitudes about externalizing behaviors changes as they age (within-person effect). (A) A younger adolescent (age 12) who exhibited greater vmPFC tracking of self-evaluations tended to endorse more permissive attitudes. However, this association flipped as they aged, such that an older adolescent (age 15) who exhibited greater vmPFC tracking of self-evaluations tended to endorse less permissive attitudes. (B) A younger adolescent (age 12) tended to endorse less permissive attitudes when they exhibited greater vmPFC tracking of perceived parent norms at the time of adolescents' choice; this effect went away as they aged (age 15). (C) vmPFC tracking of perceived peer norms at the time of adolescents' choice was not associated with adolescents' own attitudes. Simple slopes: *95% CI did not include 0.
Neural tracking of perceived parent and peer norms.
We found a significant within-person effect of vmPFC tracking of perceived parent norms and its interaction with age. As shown in Figure 4B, simple slopes analyses of this interaction effect revealed that a younger adolescent tended to report less permissive attitudes when they exhibited greater vmPFC tracking of perceived parent norms when forming their own attitudes (age 12: b = −1.13, SE = 0.30, 95% CI [−1.72, −0.54]). This effect went away as the adolescent aged (age 15: b = 0.59, SE = 0.31, 95% CI [−0.01, 1.2]). We also found a significant interaction between age and vmPFC tracking of perceived parent norms at the between-person level (Table 5), indicating that the level of vmPFC tracking of perceived parent norms moderates age differences in self attitudes between individuals. Contrary to hypotheses, there was no main effect of vmPFC tracking of perceived peer norms or its interaction with age at the within-person (Figure 4C) or between-person levels.
dACC
Neural tracking of self-evaluations.
The main effect of dACC tracking of self-evaluations and its interaction with age were nonsignificant but trended in the same direction at more liberal thresholds (Table 5).
Neural tracking of perceived parent and peer norms.
There were neither within-person effects of dACC tracking of perceived parent or peer norms nor interactions with age (Table 5). At the between-person level, there was a trending effect of dACC tracking of perceived peer norms at a more liberal threshold, but no interaction with age.
dmPFC
Neural tracking of self-evaluations.
There was a trending within-person effect of dmPFC tracking of self-evaluations at a more liberal threshold (Table 5). There were no within-person interactions with age or between-person effects.
Neural tracking of perceived parent and peer norms.
There was a trending within-person effect of dmPFC tracking of perceived parent ratings (p = .06; Table 5) and no interaction with age. There were no between-person effects.
DISCUSSION
This longitudinal fMRI study examined the neural correlates of perceived parent and peer norms that support the development of personal attitudes about externalizing behaviors from early to middle adolescence. Behaviorally, we found that adolescents perceived their peers to have more permissive norms about externalizing behaviors than their own, an effect that waned from early to middle adolescence. In contrast, adolescents perceived their parents to have less permissive norms about externalizing behaviors than their own, an effect that increased from early to middle adolescence. Consistent with the hypothesized social reorientation away from parents and toward peers across adolescence (Nelson et al., 2016), these results demonstrate that increasing self-parent discrepancies of attitudes about externalizing behaviors parallel decreasing self-peer discrepancies of attitudes from early to middle adolescence. Given the task did not elicit overt pressures to conform to the perceived norms of others, our data suggest that adolescents' own attitudes are more aligned with their perceived parent norms in the early years of adolescence but shift to be more in line with their perceived peer norms by middle adolescence.
Brain–behavior analyses revealed that adolescents' own attitudes about externalizing behaviors were predicted by the extent to which mPFC and vmPFC activity correlated with self-evaluations as well as perceived parent norms at the time of adolescents' choice. Previous studies suggest children and adults may represent the mental states of others using the same brain circuitry involved in their own choices to aid decision-making (Lim et al., 2016; Suzuki et al., 2012). Our results extend previous observations with children and adults by showing that the adolescent brain spontaneously tracks the perceived norms of parents at the same moment adolescents are making self-evaluations to ultimately inform adolescents' overall attitudes about externalizing behaviors. The vmPFC is important for representing the subjective value of information during social influence (Welborn et al., 2016), whereas the mPFC is implicated in representing self- and other-oriented knowledge, with heightened activity during adolescence (Van der Cruijsen et al., 2019; Somerville et al., 2013). Notably, this brain–behavior association was not observed in the dACC or dmPFC, which are involved in monitoring cognitive inconsistencies between self and others (Minich et al., 2023), underscoring the importance of valuation and social cognitive processes in the development of adolescent attitudes about externalizing behaviors. Thus, the recruitment of vmPFC and mPFC may reflect a shared representation between personal attitudes and perceived parent norms during adolescents' decision-making, potentially supporting developmental changes in parent influence.
Importantly, the strength of brain–behavior associations varied within a given adolescent as they aged. A younger adolescent tended to report more permissive attitudes about externalizing behaviors when they exhibited greater mPFC and vmPFC tracking of self-evaluations. However, this pattern flipped for the vmPFC as the adolescent aged, such that an older adolescent tended to report less permissive attitudes when they exhibited greater vmPFC tracking of self-evaluations. By mid-adolescence, the mPFC effect disappeared, indicating that mPFC tracking of self-evaluations was no longer associated with adolescents' own attitudes. These neural patterns may reflect the costs and benefits of relying on self-evaluations to inform personal attitudes about externalizing behaviors across adolescence, when independent decision-making is still developing (Moses-Payne et al., 2021). In early adolescence, there may be initial tradeoffs between exploiting one's limited experience over seeking advice from experienced others. However, as adolescents gain knowledge over time, greater valuation of one's knowledge may protect against external, permissive influences encountered in mid-adolescence.
Controlling for the role of self-evaluations, we found that spontaneously tracking information about perceived parent norms at the time of adolescents' own choice can buffer the endorsement of permissive attitudes about externalizing behaviors in early, but not middle, adolescence. A younger adolescent tended to report less permissive attitudes when they exhibited greater mPFC and vmPFC tracking of perceived parent norms at the time of adolescents' choice. Notably, this brain–behavior association went away by mid-adolescence. Given adolescents' own attitudes were more permissive than their perceived parent norms, our results are consistent with prior work showing that the reevaluation of personal attitudes is more likely when personal attitudes are perceived as more approving than the group norm (Perkins, 2002). In addition, we replicate and extend prior findings in young adults by demonstrating that, in early but not middle adolescents, neural tracking of parent over peer information during decisions about externalizing behaviors (including risk taking) serves as a protective factor against negative outcomes (Guassi Moreira et al., 2020, 2021). Thus, capitalizing on the influence of restrictive parent norms during decision-making in early adolescence may be crucial for slowing the development of permissive attitudes about externalizing behaviors.
Contrary to hypotheses, neural tracking of perceived peer norms was not associated with longitudinal changes in adolescent attitudes about externalizing behaviors, despite our behavioral results suggesting less divergence between adolescents' own attitudes and perceived peer norms over time. One interpretation is that the decreasing self-peer attitude discrepancy from early to middle adolescence reflects developmental changes in the representation of broader negative stereotypes of “risky” adolescents (Qu, Pomerantz, McCormick, & Telzer, 2018) rather than local peer norms outweighing parent norms. Prior work has shown that peer norms do not impact adolescent risk attitudes when parents hold strong negative views (Martino, Ellickson, & McCaffrey, 2009) or when personal attitudes are already perceived as less approving than peer group norms (Perkins, 2002). This suggests other peer factors may contribute to longitudinal changes in adolescent attitudes about externalizing behaviors.
A few limitations should be noted. First, the influence of perceived parent and peer norms may be affected by the salience of the targets or single informant perspective. However, prior research suggests known and unknown peers similarly impact adolescent risk taking (Weigard, Chein, Albert, Smith, & Steinberg, 2014) and that risk behaviors are more strongly related to perceived than actual norms (Perkins et al., 2019). Future studies should explore whether similar parent-over-peer preferences in the neural tracking of perceived norms hold when adolescents make choices involving a closer peer (e.g., best friend) relative to an unknown peer. Second, investigating the moderating roles of parent and peer relationship quality, as well as adolescents' actual engagement in externalizing behaviors, would provide valuable added insights into the co-development among personal attitudes and the perceived norms of parents and peers. Finally, the study focused on midline cortical regions involved in self–other processing, and further research could examine the involvement of other brain regions or functional connectivity patterns on longitudinal changes in adolescent attitudes about externalizing behaviors.
The present study reveals the neural correlates of adolescents' perceptions of parent and peer norms and their role in supporting longitudinal changes in personal attitudes about externalizing behaviors. Findings reveal how perceived parent and peer norms change in parallel with personal attitudes about externalizing behaviors from early to middle adolescence and underscore the importance of spontaneous neural tracking of perceived parent norms during self-evaluations for buffering personal attitudes, particularly in early adolescence.