Abstract
Question: What functional brain connections are associated with alcohol use risk in adolescents, and are these connections different for male vs female adolescents? Findings: This cohort study of 1359 adolescents identified common and distinct neurobiological substrates of alcohol use as well as sex divergence in model accuracies, such that models predicting female risk consistently outperformed models predicting male risk. Meaning: The networks identified in this study and subsequently externally validated represent candidate targets for future prevention and intervention efforts.
Introduction
Adolescence is a critical time for transitions from alcohol misuse and experimentation to disorder due to ongoing brain development during this time. This period is therefore considered an essential point for identification of brain circuits contributing to problems later in life, a phenomenon referred to as preaddiction. Independent of future adult drinking, high-risk alcohol use during adolescence is itself an adverse health behavior, associated with multiple negative sequelae, and represents a leading cause of disability and mortality. By identifying brain mechanisms associated with early risk, we aim to facilitate the future development of novel prevention and intervention approaches to help mitigate problematic alcohol use.
Contemporary theories of alcohol use risk emphasize the dual roles of top-down inhibitory systems involving prefrontal cortical structures and bottom-up subcortical reward systems. However, studies of the brain’s functional connectome through network neuroscience indicate that typical brain maturation involves complex functional reorganization of networks instead of simple linear maturation of individual regions. These insights have transformed understanding of basic neurodevelopment but are not well characterized within the context of alcohol use risk behaviors.
Sex differences in neurodevelopment have been widely noted, with female individuals displaying accelerated developmental trajectories relative to male individuals. Sex differences in alcohol use during adolescence are also commonly reported. Therefore, this study aimed to identify neural networks conferring vulnerability for alcohol use in adolescents with specific consideration of sex differences. We adopted a multivariate predictive modeling framework to identify and cross-validate (ie, test in novel data) brain networks associated with current and future alcohol use risk in a large sample (approximately 1500) of adolescents using neuroimaging data acquired during performance of different cognitive tasks relevant to current neurodevelopmental models: that is, a reward-processing task and an inhibitory control task. Based on contemporary theories of neurodevelopment (eAppendix in Supplement 1), we anticipated that networks identified during reward-related processes would be most relevant for predicting alcohol use behaviors in younger adolescents (approximately 14 years) vs older adolescents (approximately 19 years), and that networks identified during inhibitory-related processes would be most relevant for predicting alcohol use behaviors in older vs younger adolescents.
Methods
Overview
The IMAGEN consortium collected alcohol and neuroimaging data across 8 sites. Ethics committees at each site approved the study. A parent or guardian provided written consent, and adolescents provided verbal assent. The study followed the Strengthening the Reporting of Observational Studies in Epidemiology (STROBE) reporting guideline. Figure 1 provides a schematic overview of primary data collection and analysis steps. Baseline data were acquired at age 14 years. Follow-up data were acquired at age 19 years. Analyses focused on predicting alcohol use risk at age 19 years. Data acquired at baseline (age 14 years) were used to identify networks associated with future alcohol use risk, and data acquired at follow-up (age 19 years) were used to identify networks associated with current alcohol use risk.
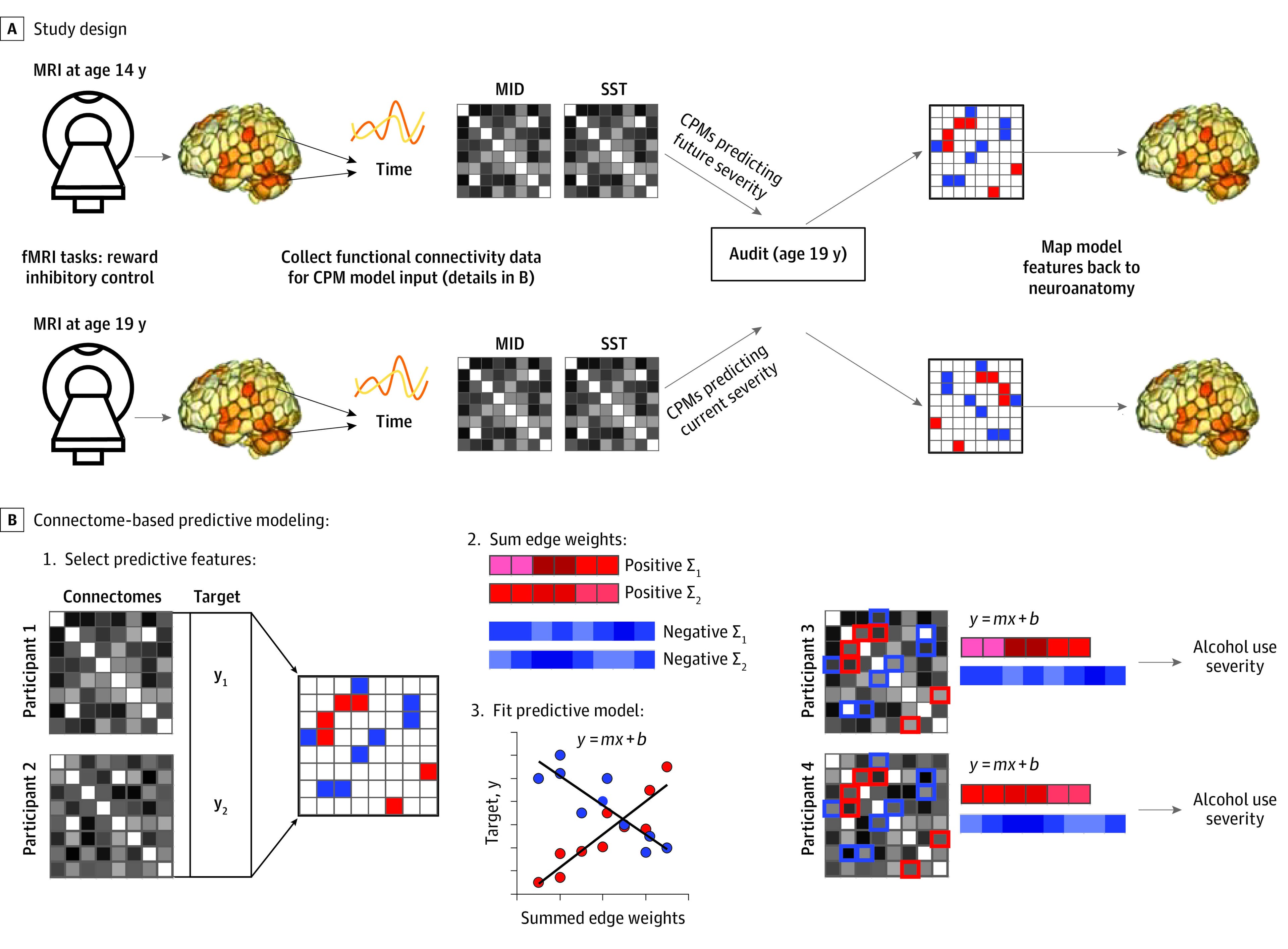
Neuroimaging data were processed using a validated functional connectivity pipeline (eMethods in Supplement 1) to generate individual participant functional connectivity matrices (hereafter referred to as connectomes). Connectomes provide a multivariate summary of an individual’s unique pattern of functional brain organization. While connectomes are relatively distinct across individuals (ie, able to uniquely identify individuals, a process referred to as neural fingerprinting), an individual’s connectome also varies as a function of cognitive task performance, or brain state. Moreover, recent work demonstrates that brain state manipulation (via cognitive task performance vs resting state) is optimal for identifying core individual difference factors, including predictive neuromarkers of substance use in adults. Our analyses focus on connectomes derived from neuroimaging data acquired during both reward and inhibitory tasks (Figure 1A). Connectomes were entered into connectome-based predictive models (CPMs) (Figure 1B) to identify networks associated with alcohol misuse.
Participants
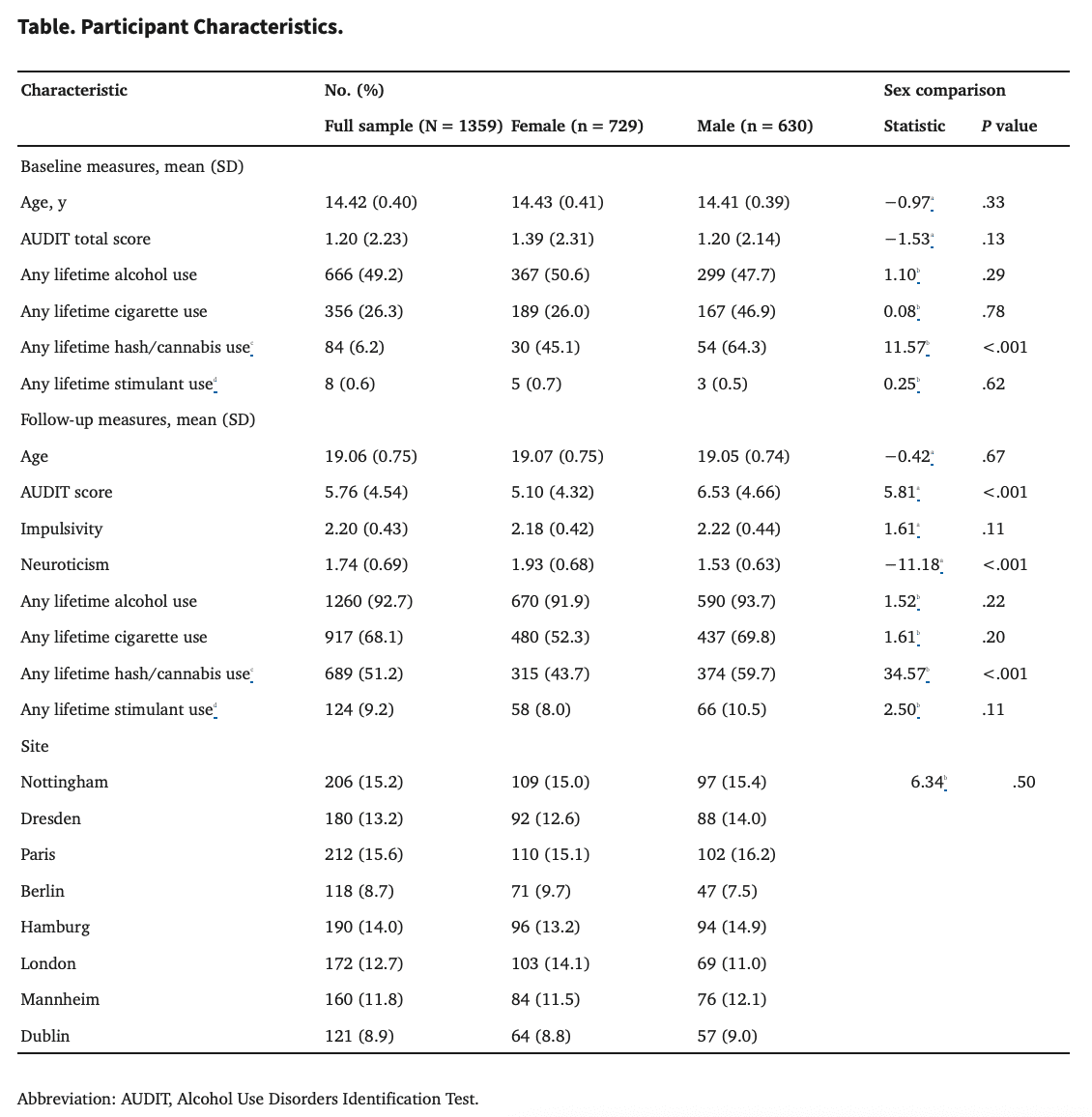
Demographic characteristics are provided in the Table. An independent sample of US college students (n = 114; age 17-23 years at scan, mean [SD] age, 18.42 [0.77] years) was used as an external replication sample (in Supplement 1).
Alcohol Use
Alcohol use was assessed using the Alcohol Use Disorders Identification Test (AUDIT), a well-validated self-report measure of alcohol use risk with total scores ranging from 0 to 40. AUDIT scores at age 14 years were low with relatively little variance across the sample (mean [SD] score, 1.36 [2.42]) (eFigure 1 in Supplement 1). Variance in AUDIT scores at age 19 years was higher (mean [SD] score, 5.53 [4.47]), and scores increased significantly from age 14 years to 19 years (t870 = 28.94; P < .001). The Table and eFigure 2 in Supplement 1 present details on other substance use.
Neuroimaging
Functional magnetic resonance imaging (fMRI) data were collected during a reward task (monetary incentive delay) and during an inhibitory control task (stop signal). Details on tasks, preprocessing, and connectivity analyses are in the eMethods and eFigures 3 and 4 in Supplement 1.
Connectome-Based Predictive Modeling
As shown in Figure 1B, CPM comprises the following steps: (1) feature selection, wherein regression is used to identify connectome features associated with a behavioral variable of interest (here, alcohol use severity, using a feature selection threshold of P < .05) in a training data set; (2) feature reduction, in which identified connections are summed to create a summary value for each individual in the training data set; (3) model building, wherein summary scores (independent variable) are linearly associated with the behavioral variable (dependent variable); (4) model application, in which resultant linear models are applied to novel connectomes in a testing data set to generate predictions; and (5) model evaluation, in which the predictive ability of the model is evaluated based on the correspondence between model-predicted and actual alcohol use risk scores. Unlike correlation or regression, CPM with built-in cross-validation protects against overfitting by testing the strength of the association in previously unseen data (step 5), increasing the likelihood of replication in future studies and thus applicability to other clinical samples.
Connectomes for each task and time point were entered into CPMs using a nested cross-validation scheme, wherein 1 test site was left out, and 10-fold cross-validation was used for the left-in training sites. In this approach, data from a single site is left out. The predictive model is then generated using the remaining data from the other 7 sites. Model accuracy is then tested in the left-out site. This approach is the current gold standard for multisite neuroimaging data.
Both sex-agnostic (both sexes combined) and sex-specific (female and male separately) models were conducted. Model results were Bonferroni corrected across the number of models run at each time point and considered significant at 2-tailed α < .008, as determined via permutation testing.
Network Anatomy and Virtual Lesioning
Consistent with best practice recommendations, we summarized the anatomy of identified networks associated with alcohol use risk via their spatial overlap with well-established large-scale neural networks (eFigure 5 in Supplement 1), hereafter referred to as canonical neural networks. Further, consistent with recommendations to maximize mechanistic insight, we used a post hoc virtual lesioning approach in which connections overlapping with all but a single canonical network (eg, default mode) were virtually lesioned, and the model was rerun. By doing this for each canonical network separately, we were able to determine the relative predictive accuracy, or feature importance, of connections within each of these established networks. Network masks are available for download.
Results
Cross-Validated Brain-Behavior Models
Neural Networks Associated With Future Alcohol Use Risk
Among 1359 individuals in the study, the mean (SD) age was 14.42 (0.40) years, and 729 individuals (54%) were female. Analyses of baseline data (age 14 years) found sex divergence in the predictive accuracy of CPMs assessing future alcohol use severity. Specifically, sex agnostic models (reward ρ, 0.17; total edges, 1575; P < .001; inhibition ρ, 0.22; total edges, 1640; P < .001) and female-only models (reward ρ, 0.29; total edges, 3150; P < .001; inhibition ρ, 0.26; total edges, 2119; P < .001) were successful in predicting future alcohol use severity at age 19 years, but male-only models were not (reward ρ, 0.04; total edges, 359; P = .43; inhibition ρ, 0.11; total edges, 391; P = .02). The same pattern of results emerged in post hoc analyses controlling for baseline alcohol use severity and residual motion (eResults and eTable 1 in Supplement 1). Comparisons of model accuracies using Fisher r to zconfirmed that the female-only models significantly outperformed male-only models (reward z = 4.0; P < .001; inhibition z = 2.47; P = .01).
Neural Networks Associated With Current Alcohol Use Risk
Analyses of follow-up data (age 19 years) further indicated sex divergence in the predictive accuracy of CPMs assessing current alcohol use severity. Specifically, both sex-agnostic models (reward ρ, 0.17; total edges, 2904; P < .001; inhibition ρ, 0.27; total edges, 2538; P < .001) and female-only models (reward ρ, 0.29; total edges, 2606; P < .001; inhibition ρ, 0.27; total edges, 2134; P < .001) were successful in predicting current severity across both task types. In contrast, male-only models were successful in predicting current severity using connectivity data acquired during inhibitory task performance (ρ, 0.18; total edges, 622; P < .001) but not using data acquired during reward task performance (ρ, 0.03; total edges, 906; P = 0.44), indicating task specificity of alcohol risk models in male individuals only.
The same pattern of results emerged in post hoc analyses controlling for baseline alcohol use severity and residual motion (eTable 2 in Supplement 1). Comparisons of model accuracies indicated that the female-only model generated from reward data significantly outperformed the male-only model generated from reward data (z = 4.4; P < .001). There were no significant differences in predictive model performances between female-only and male-only models generated from inhibitory data (z = 1.67; P = .10).
Anatomy of Alcohol-Risk Networks
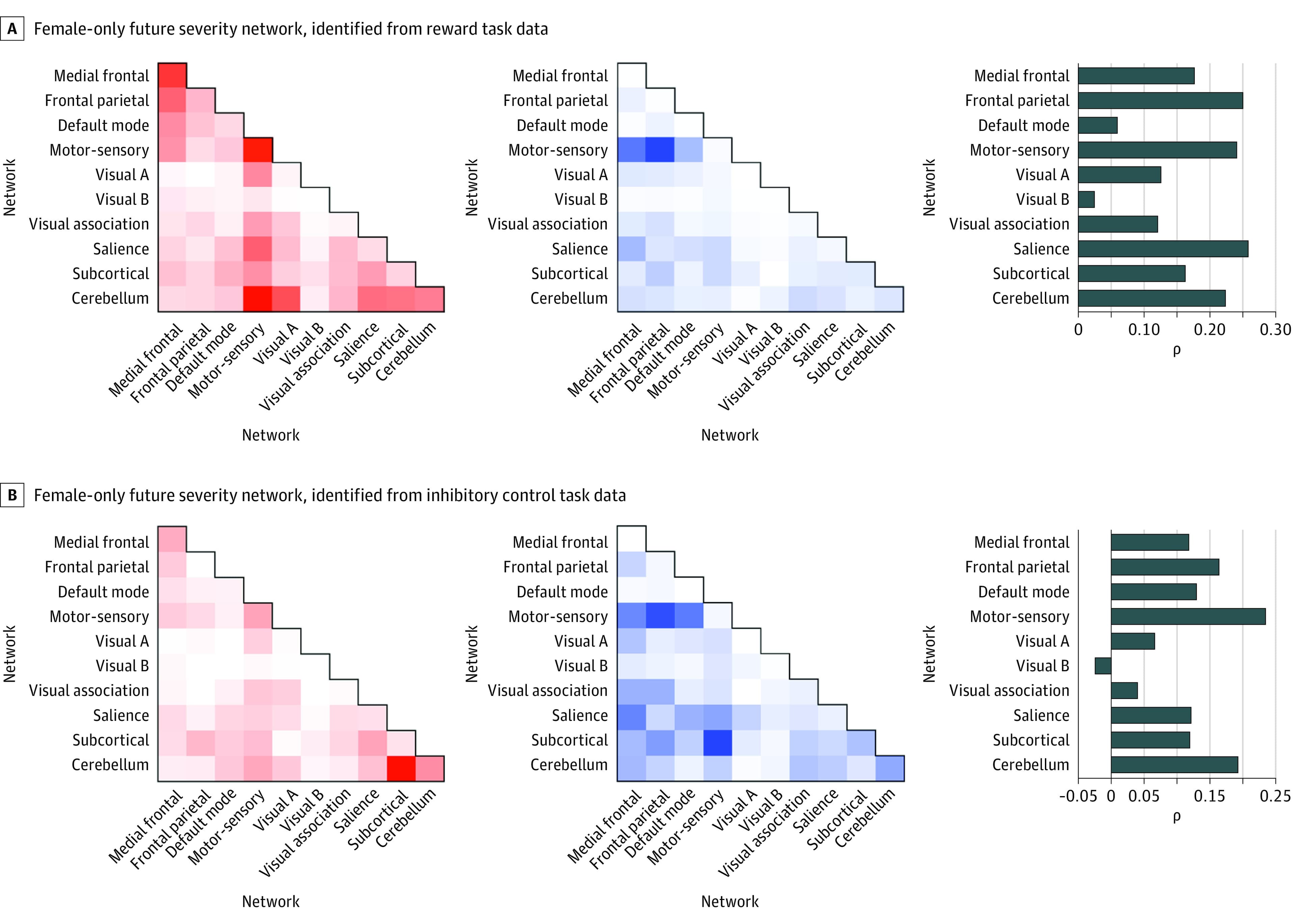
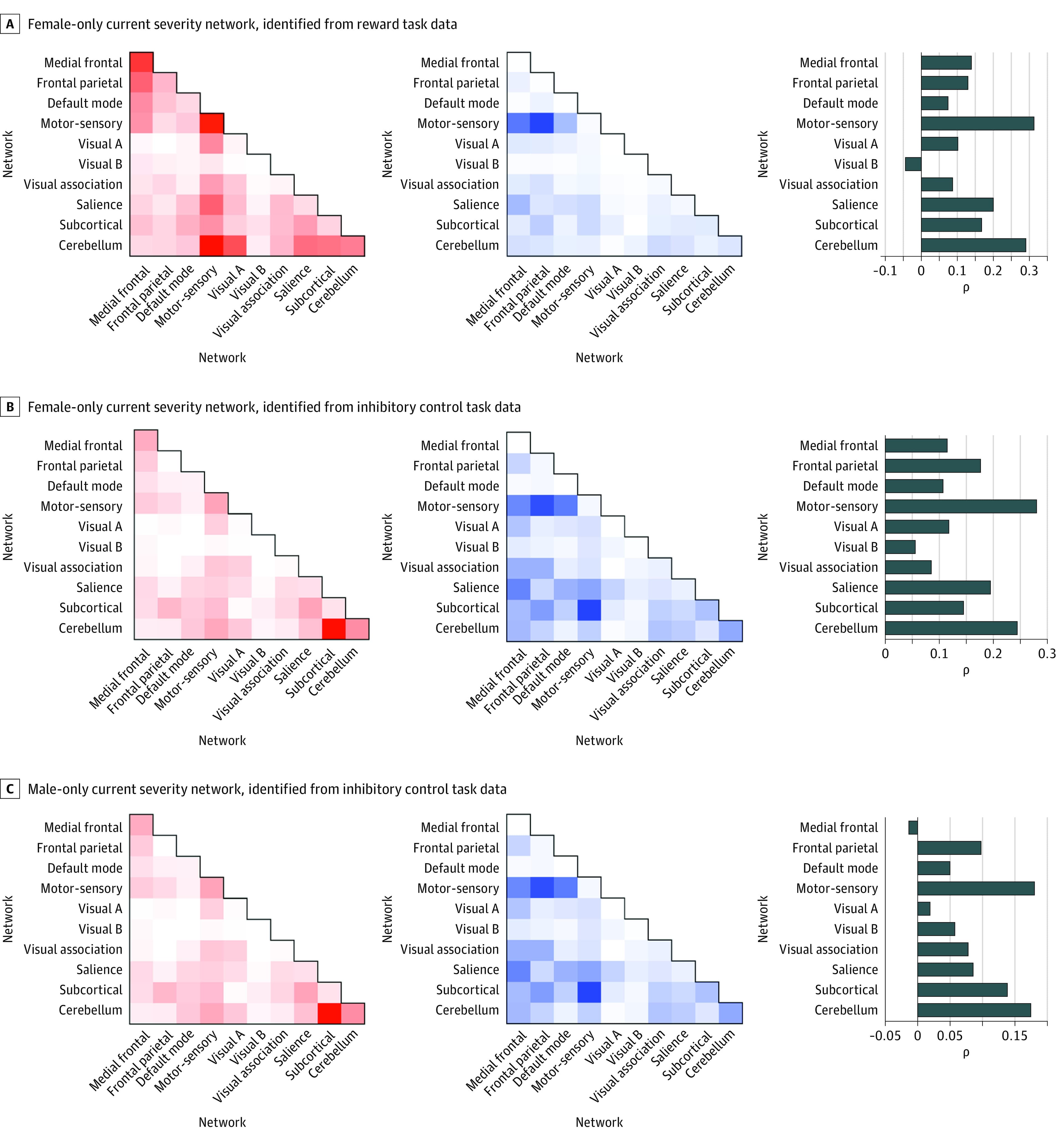
Figures 2 and 3 show the anatomy of the identified networks as a function of spatial overlap with canonical neural networks. Anatomy is summarized separately for positive and negative predictive connections. Positive predictive connections are those for which increased connectivity positively predicted alcohol use risk. Negative predictive connections are those for which decreased connectivity positively predicted alcohol use risk. By definition, positive and negative connections cannot overlap directly (as a single connection cannot be both a positive and negative predictor). Nonetheless, as summarized in Figures 2 and 3, positive and negative predictive connections may include connections within and between similar large-scale canonical neural networks. Figures 2 and 3 also show virtual lesioning results corresponding to the relative predictive accuracy or feature importance of connections.
Neural Networks Associated With Future Alcohol Use Risk
As shown in Figure 2, neural networks associated with future alcohol use using either reward or inhibitory control task data were similar among female adolescents, indicating the relevance of both processes in conferring vulnerability for alcohol use risk. For both tasks, positive connections included a high degree of within-network medial frontal, motor-sensory, and cerebellar connections, as well as substantial between-network connections with the cerebellar and subcortical networks. The network identified using reward data was further characterized by between-network connections within the medial frontal and motor-sensory networks. For both tasks, negative connections included a high degree of between-network motor-sensory connections.
Post hoc virtual lesioning indicated that networks with the highest individual feature weights—that is, amount of variance in future alcohol use associated with connectivity within a given canonical network—included frontoparietal, salience, motor-sensory, and cerebellar networks.
Neural Networks Associated With Current Alcohol Use Risk
As shown in Figure 3, female-only positive networks generated from both tasks were dominated by motor-sensory, cerebellar, subcortical, and salience network connections, whereas negative networks were largely characterized by motor-sensory, salience, and subcortical connections. Although the male-only networks identified using inhibitory data were sparser than the female networks, they featured connections within and between similar networks: the positive male-only network was dominated by connections between cerebellar and subcortical networks as well as by within-network cerebellar connections; the negative male-only network contained a high degree of cerebellar, subcortical, salience, and medial frontal connections as well as a high degree of within-network subcortical connections.
For female adolescents, post hoc virtual lesioning indicated that networks with the highest individual feature weights—that is, amount of variance in current alcohol use associated with connectivity within a given canonical network—included salience, motor-sensory, and cerebellar networks. For male adolescents, virtual lesioning indicated that networks associated with the highest amount of variance in current alcohol use included motor-sensory, cerebellar, and subcortical regions.
Network Change Over Time
For successful models, individual participant network summary scores were created as the sum of connectivity strengths within positive and negative networks and entered into paired t tests to characterize the development of the identified networks over time. Among female adolescents, connectivity within reward and inhibitory control networks predictive of future alcohol use significantly increased from age 14 years to age 19 years (reward t539 = −7.09; P < .001; inhibition t559 = −4.68; P < .001). Among both female and male adolescents, connectivity within inhibitory control networks predictive of current use also significantly increased from age 14 to 19 (female t = −5.61(df = 559), P < .001; males: t468 = −5.87; P < .001), while changes in connectivity within the reward network predictive of current use in female adolescents from age 14 years to age 19 years did not reach significance (t539 = −1.77; P = .08).
Network Overlap Across Sexes
Despite significant differences in predictive accuracies, networks identified as associated with alcohol use behaviors were relatively consistent across male and female adolescents. Consistent with this, cross-sex analyses demonstrated that models developed in female adolescents at age 19 years generalized to predict alcohol use in male adolescents at age 19 years (reward: ρ, 0.14; P = .001; inhibition: ρ, 0.15; P < .001), and that models developed in male adolescents at age 19 years generalized to predict alcohol use in female adolescents at age 19 years for inhibitory, but not reward, task data (reward: ρ, 0.07; P = .08 inhibition: ρ, 0.26; P < .001).
Specificity and Generalizability
Specificity Analyses
To determine the specificity of the identified anatomical connections to alcohol use, we repeated CPMs while controlling for variation in impulsivity and neuroticism. For all models, predictive accuracies were robust to these factors, indicating that the identified connections were specific to alcohol use and that model accuracies were not driven by a more general latent trait personality factor (eTables 3 and 4 Supplement 1).
As shown in the Table, substance use rates were low at age 14 years but were higher at age 19 years (eg, approximately 64% lifetime cigarette use). Post hoc analyses indicated significant associations between substance use and connectivity within the male-only inhibition model such that connectivity was greater among males with nonalcohol substance use. Follow-up analyses indicated that associations between alcohol use and connectivity remained significant within each group (substance-use positive and substance-use negative) separately (eTables 3-5 in Supplement 1).
Generalizability
The ultimate test of statistical reliability is whether a model produces consistent results in novel individuals and settings. For this reason, in our analyses above, we adopted a rigorous leave-one-site-out cross-validation scheme, in which models were tested in previously unseen testing sites. As a further test of the robustness of the effects, we sought to determine whether the identified neural networks would also be associated with alcohol use risk in an entirely separate sample of 114 adolescents from the Brain and Alcohol Research in College Students (BARCS) study who were recruited independently from the IMAGEN consortium (eResults in Supplement 1).
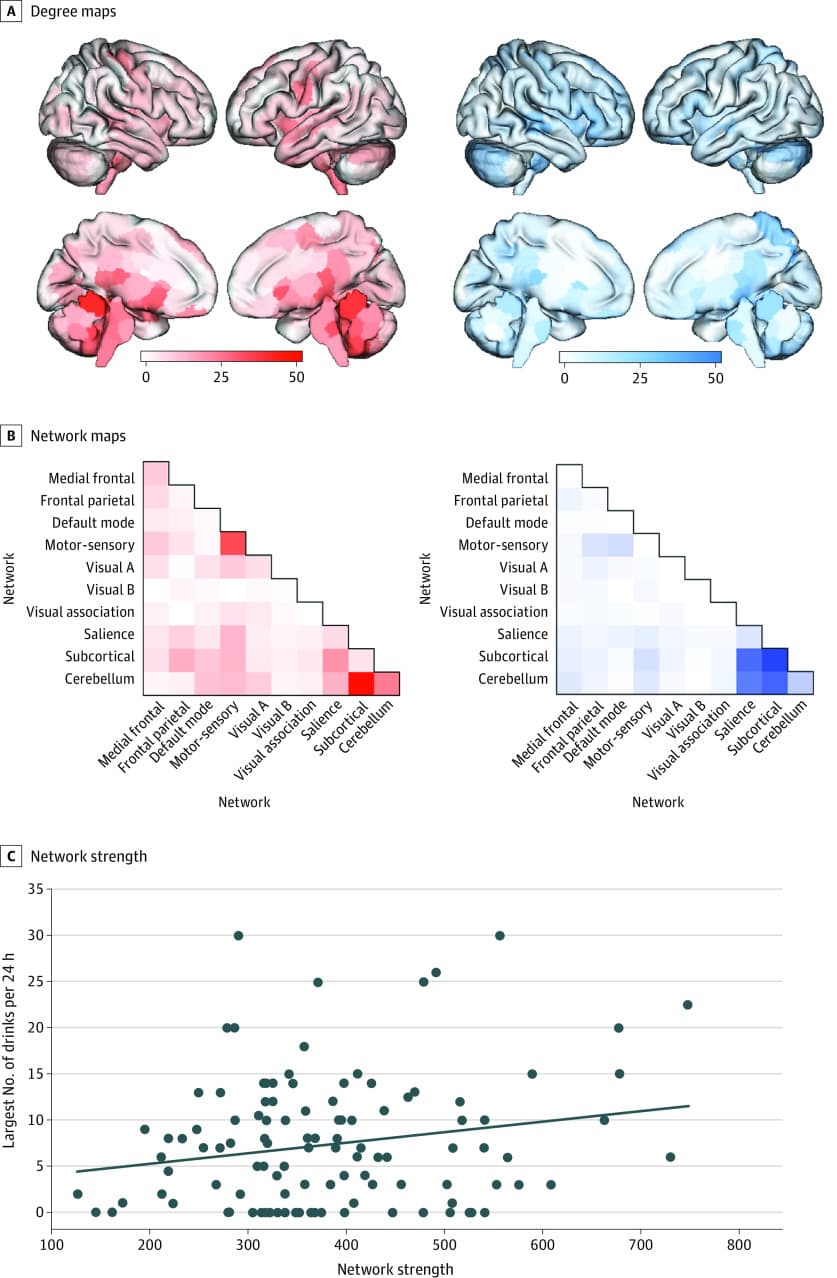
BARCS differed from IMAGEN in multiple ways, including a different within-scanner inhibitory control task (go/no-go task), different imaging acquisition parameters, a smaller sample size, recruitment from the US (vs Europe), and different alcohol use risk assessments. Despite these differences, connections identified in IMAGEN as associated with current alcohol use risk replicated to predict risky alcohol use (in this case, defined as the largest number of alcoholic drinks consumed in 24 hours) in this external test sample (ρ, 0.20; total edges, 113; P = .03) (Figure 4). Additional details and analyses related to BARCS are in the eResults in Supplement 1.
Discussion
This study leveraged data from the IMAGEN consortium to identify neural networks associated with alcohol use risk using an advanced connectome-based approach. Using data from 2 cognitive tasks with relevance to adolescent risk behaviors—a reward task and an inhibitory control task—we identified both common and distinct neurobiological substrates of early alcohol use risk among female and male adolescents, as well as sex divergence in model accuracies. For female adolescents, models generated using neuroimaging data from both task types were successful in identifying reliable neural signatures of future and current alcohol use risk. In contrast, for male adolescents, only the model generated using inhibitory control task data was successful in predicting current alcohol use risk, and no reliable signature of future risk was identified (ie, brain features identified in training data did not predict alcohol use in previously unseen testing data). Despite these differences, connections identified as commonly associated with current alcohol use risk across male and female adolescents not only generalized to held-out testing sites within IMAGEN, but also predicted alcohol behaviors in an entirely independent cohort of adolescents. Thus, the identified alcohol use risk network may be considered as a robust neuromarker that may be targeted in future prevention and intervention efforts.
Among female adolescents, identified connections included subcomponents of multiple well-established resting state networks, including medial frontal, frontoparietal, salience, motor-sensory, and cerebellar networks. Connectivity within these networks generally increased from age years 14 to age 19 years, concurrent with increases in alcohol use. In addition, despite similar features overall, the relative predictive weights of network subcomponents associated with future (age 14 years) vs current (age 19 years) alcohol use risk varied over time, such that medial frontal and salience networks were primarily associated with future alcohol use risk, whereas cerebellar and motor-sensory networks emerged as primarily associated with current alcohol use risk. These findings are consistent with recent data demonstrating altered cerebellar growth trajectories among adolescents with heavy alcohol consumption as well as with much broader literature implicating cerebellar and motor systems in substance-use pathophysiology (eDiscussion in Supplement 1). Together, these data suggest that targeting prefrontal cortical regions may be particularly relevant to prevention efforts in younger adolescents and that targeting cerebellar and motor-sensory regions may be more relevant for intervention efforts in older adolescents who have already initiated drinking. Consistent with this, network connectivity at age 19 years generalized to predict alcohol use in our external sample of older young adults.
Inhibitory control (vs reward) brain states were more relevant for predicting alcohol use in older adolescents; however, this effect was specific to male adolescents. This finding is consistent with work demonstrating that the accuracy of generalizable brain-behavior models of core individual difference factors (eg, intelligence) differs as a function of both sex and brain state, but for the first time to our knowledge demonstrates similar sex specificity within the context of alcohol use risk. Together, these data suggest that interventions focusing on inhibitory control processes may be particularly effective in combating current alcohol use risk in male adolescents but that both inhibitory and reward-related processes are likely of relevance to current alcohol use behaviors in female adolescents. Sex-specific tailoring of interventions has been proposed previously but is seldom—if ever—grounded in neurobiology. By identifying robust markers of risky alcohol use during development, we hope to support the development of such neurobiologically informed prevention and intervention efforts.
Strengths and Limitations
This study has several strengths, including its unique longitudinal sample and the use of a wholly data-driven approach with the incorporation of rigorous leave-one-site-out internal validation and external validation using an independent sample. This study also has several limitations, including the absence of a consistent measure of alcohol use severity across internal and external validation samples and the absence of more frequently acquired neuroimaging data that could be used to track trajectories. Further, our external replication sample was primarily White European, so it remains to be seen whether these predictive networks will generalize to more diverse populations.
Conclusion
These data provide a connectome-wide assessment of neural networks subserving alcohol use risk and identify a dimensional neural signature of alcohol use risk in adolescents. External validation supported these findings.